科学家用非转基因水稻和转Z基因水稻为实验材料,分别喷施蒸馏水、一定浓度的寡霉素和一定浓度的NaHSO3,24h后进行8h的干旱胁迫处理,测得胁迫前后水稻光合速率如下表所示。已知寡霉素抑制光合作用和细胞呼吸中ATP合酶的活性。回答下列问题:
(1)ATP合酶能催化合成ATP的作用机理是____________________ 。寡霉素在细胞呼吸和光合作用过程中的主要作用部位分别是___________ 和___________ 。
(2)实验中实际测得的光合速率是净光合速率,若要获得水稻的总光合速率,还需要__________ 。
(3)分析表中数据可知,喷施NaHSO3能够____________________ ;植物吸收的NaHSO3中的S元素可用于合成___________ 等(至少答出1种)生物大分子。
(4)研究发现,一种名为Steptomyces-d-m的土壤放线菌能产生大量的寡霉素,推测___________ (填“非转基因”或“转Z基因”)水稻更能适应富含Steptomyces-d-m的土壤,判断的依据是_____________________________ 。
实验材料 | 实验处理 | 光合速率(umolCO2•m-2•s-1) | ||
胁迫前 | 胁迫后 | |||
非转基因水稻 | 1组 | 喷施蒸馏水 | 16.31 | 5.93 |
2组 | 喷施一定浓度的寡霉素 | 11.27 | 4.51 | |
3组 | 喷施一定浓度的NaHSO3 | 18.29 | 9.43 | |
转Z基因水稻 | 4组 | 喷施蒸馏水 | 18.37 | 12.14 |
5组 | 喷施一定浓度的寡霉素 | 16.03 | 10.98 | |
6组 | 喷施一定浓度的NaHSO3 | 20.34 | 16.81 | |
(2)实验中实际测得的光合速率是净光合速率,若要获得水稻的总光合速率,还需要
(3)分析表中数据可知,喷施NaHSO3能够
(4)研究发现,一种名为Steptomyces-d-m的土壤放线菌能产生大量的寡霉素,推测
23-24高三上·江西宜春·期末 查看更多[6]
江西省宜春市袁州区一中2023-2024学年高三1月期末生物试题湖南省九师联盟2023~2024学年高三上学期期末生物试题山西省忻州市2023-2024学年高三1月期末理综生物试题(已下线)2024年新课标高考生物真题吉林卷变式题19-21(已下线)2024年新课标高考生物真题辽宁卷变式题19-21(已下线)2024年新课标高考生物真题黑龙江卷变式题19-21
更新时间:2024/02/03 08:37:14
|
相似题推荐
非选择题-解答题
|
适中
(0.65)
名校
解题方法
【推荐1】沙棘耐干旱、耐盐碱、抗风沙能力强,被广泛用于水土保持。科研人员利用“ 间 隙光”(光照 20 秒、黑暗 20 秒交替进行)处理沙棘叶肉细胞一段时间,部分实验结果如图 所示。回答下列问题:
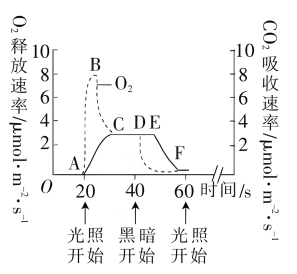
(1)沙棘叶肉细胞产生 NADH 的部位有______ ,消耗 NADPH 的部位有______ 。
(2)B 点时叶肉细胞的光合速率______ (填“大于”、“等于”或“小于”)呼吸速率。黑暗 开始后实线 DEF 段与虚线 DF 段不重合的原因是______ 。
(3)研究发现,沙棘细胞内可溶性蛋白、可溶性糖等物质的含量较多,其意义是______ , 从而有利于沙棘适应干旱、盐碱的生存环境。
(1)沙棘叶肉细胞产生 NADH 的部位有
(2)B 点时叶肉细胞的光合速率
(3)研究发现,沙棘细胞内可溶性蛋白、可溶性糖等物质的含量较多,其意义是
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐2】一种突变体大豆,其叶绿素含量仅是野生型大豆的51%,但固定CO2酶的活性显著高于野生型。下图1表示大豆叶肉细胞中相关物质变化过程,图2显示两种大豆在适宜温度、不同光照强度下的CO2吸收速率。请回答相关问题:

(2)在图2中P点时,给野生型大豆提供H218O,则18O最先出现在图1中的物质
(3)分析图2可知,当光照强度低于Q时,野生型的光反应强度
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐3】某生物研究小组在密闭恒温玻璃温室内进行植物栽培试验,连续48 h测定温室内CO2浓度及植物吸收CO2速率,得到图所示曲线(整个过程呼吸作用强度恒定),请回答下列相关问题:
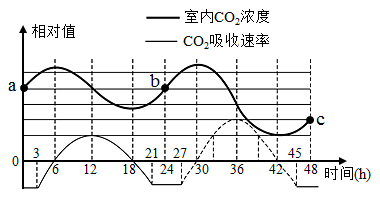
(1)实验开始阶段的0~3小时,叶肉细胞产生ATP的场所有________________ ,图中植物呼吸速率与光合速率相等的时间点有________ 个。
(2)6时CO2在叶肉细胞内的移动方向是________________ ,由12时到18时,叶绿体内三碳化合物含量变化是________ 。
(3)叶绿体利用CO2速率最大的时刻是________ h时。
(4)如果使用相同强度绿光进行实验,c点的位置将________ (填上移下移或不变),原因是__________________________________________________ 。
(1)实验开始阶段的0~3小时,叶肉细胞产生ATP的场所有
(2)6时CO2在叶肉细胞内的移动方向是
(3)叶绿体利用CO2速率最大的时刻是
(4)如果使用相同强度绿光进行实验,c点的位置将
您最近一年使用:0次
非选择题-实验题
|
适中
(0.65)
【推荐1】植物细胞中的叶绿体在光照条件下消耗一定量的 O2, 这属于植物细胞光呼吸的重要环节,如图表示光合作用和光呼吸的部分过程,①~⑤表示生理过程,X、Y、Z代表物质。请回答:
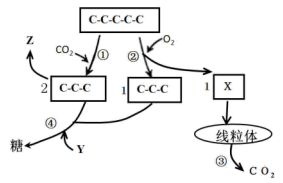
(1)图中X表示的物质是__________ 碳化合物,经过程③__________ (填生理过程名称)产生 CO2。
(2)光呼吸也必须在有光的条件下发生,因为在黑暗条件下, C-C-C不能经还原再生成____________ 。
(3)催化过程①和②的均是Rubisco 酶,CO2和 O2会竞争酶的同一活性位点,结合形成 。当细胞中 CO2/O2偏低时,会__________ (填“促进”或“抑制”)②过程。
(4)图中过程④需要光反应提供物质Y,某科学小组用来自新鲜菠菜的类囊体提取液和光合色素提取液开展以下实验,如下表:
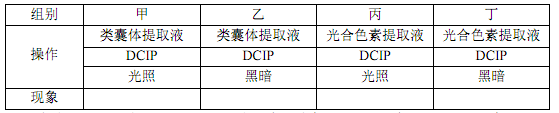
注:染料 DCIP(二氯酚靛酚)溶液的氧化态呈蓝色,被还原后会出现明显的颜色变化。
上表__________ 组会出现明显的颜色变化。氧化型 DCIP 的作用类似于光反应过程中的__________ (填物质),它既可利用于颜色反应,被还原后还可作为________________ 。
(1)图中X表示的物质是
(2)光呼吸也必须在有光的条件下发生,因为在黑暗条件下, C-C-C不能经还原再生成
(3)催化过程①和②的均是Rubisco 酶,CO2和 O2会竞争酶的同一活性位点,结合形成 。当细胞中 CO2/O2偏低时,会
(4)图中过程④需要光反应提供物质Y,某科学小组用来自新鲜菠菜的类囊体提取液和光合色素提取液开展以下实验,如下表:
注:染料 DCIP(二氯酚靛酚)溶液的氧化态呈蓝色,被还原后会出现明显的颜色变化。
上表
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐2】大豆与根瘤菌是互利共生关系,下图所示为大豆叶片及根瘤中部分物质的代谢、运输途径。回答下列问题:
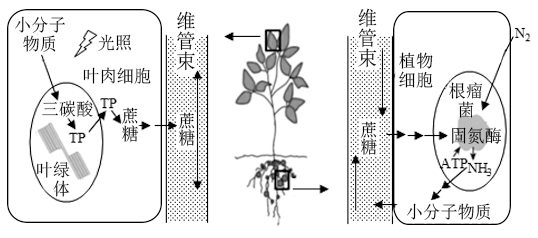
(1)叶绿体中的光合色素分布在________ 上,这些色素吸收、传递、________ 可见光的光能,其中叶绿素a主要吸收__________ 。
(2)直接固定CO2的化学物质是________ ,固定产物三碳酸被还原后产生TP——_________ (填中文名)。催化以上一系列反应的相关酶存在于__________ 。运出叶绿体的TP除图中去路外,还可以_________ 。据图可知,叶肉细胞合成的蔗糖长距离运输后__________ (填“能”或“不能”)被植物的其他细胞吸收利用。
(3)N2经________ 方式通过植物细胞与根瘤菌的质膜进入根瘤菌细胞内,被固定后产生的NH3可用于________ 、________ 等小分子物质的合成,这些小分子物质通过________ 反应形成高分子化合物控制、承载细胞的生命活动。
(4)CO2和N2转化成有机物都需要消耗大量ATP。叶绿体合成ATP的能量来自________ 过程;根瘤中合成ATP的能量主要源于_________ 的分解。
(1)叶绿体中的光合色素分布在
(2)直接固定CO2的化学物质是
(3)N2经
(4)CO2和N2转化成有机物都需要消耗大量ATP。叶绿体合成ATP的能量来自
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
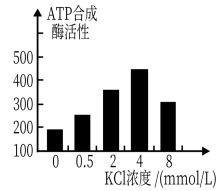
【推荐3】为研究氯化钾(KCl)对烟草光合作用和ATP合成酶活性的影响,科研人员采用水培漂浮育苗方法进行了有关实验,实验过程中,每7天更换一次培养液,实验结果如下所示:
回答下列问题:
(1)实验过程中,每7天更换一次培养液的目的是____________ 。
(2)由表可知:导致4 mmol/L KCl溶液条件下烟草净光合速率小于2 mmol/L KCl溶液条件下烟草净光合速率的因素不是CO2浓度,其理由是______________ 。
(3)ATP合成酶参与光合作用的_________ 阶段。在4 mmol/L的KCl溶液中,ATP合成酶活性最高,这主要有利于光合作用的_________ (填“物质转化”或“能量转换”。但此浓度下植物净光合速率却不是最高,其原因可能是___________________________ 。
氯化钾浓度(mmol/L) | 气孔导度[mmol/(m2•s)] | 净光合速率[pmol/(m2•s)] |
0 | 0.18 | 4.52 |
0.5 | 0.19 | 6.96 |
2 | 0.26 | 9.30 |
4 | 0.28 | 7.16 |
8 | 0.25 | 5.71 |
(1)实验过程中,每7天更换一次培养液的目的是
(2)由表可知:导致4 mmol/L KCl溶液条件下烟草净光合速率小于2 mmol/L KCl溶液条件下烟草净光合速率的因素不是CO2浓度,其理由是
(3)ATP合成酶参与光合作用的
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐1】“海水稻”是指能够在盐碱地上生长的水稻。研究者将某品种海水稻随机均分成甲、乙两组,分别在盐碱地和非盐碱地中种植一段时间,测定两组植株净光合速率随光照强度的变化情况。结果如下表所示。请回答下列问题:
(1)海水稻进行光合作用时,绿叶从外界吸收的CO2与C5结合形成C3,这个过程称作______ 。C3接受光反应阶段生成的______ ,形成______ 。
(2)光照强度0~600Lx,限制光合速率的因素主要是______ 。光照强度为600Lx时,两组植株的光合速率______ (填“甲>乙”“甲=乙”或“甲<乙”),判断依据是______ 。
(3)光照强度由1600Lx上升至3200Lx时,乙组光合速率继续增大,而甲组不再增大,推测可能是种植在盐碱地中的海水稻光合色素含量减少造成的,为验证该推测,请简要写出实验思路:______ 。
光照强度(Lx) | 0 | 200 | 400 | 600 | 800 | 1600 | 3200 | 6400 | |
净光合速率[μmmolCO2/(m2·a)] | 甲组 | -3 | 0 | 2 | 6 | 10 | 12 | 12 | 12 |
乙组 | -10 | -5 | 0 | 6 | 12 | 18 | 20 | 20 | |
(2)光照强度0~600Lx,限制光合速率的因素主要是
(3)光照强度由1600Lx上升至3200Lx时,乙组光合速率继续增大,而甲组不再增大,推测可能是种植在盐碱地中的海水稻光合色素含量减少造成的,为验证该推测,请简要写出实验思路:
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐2】夏季晴朗无云的某天,某种绿色植物光合作用强度变化曲线如图所示。请回答下列问题:
(1)在A点时植物细胞中会产生ATP的场所是_________
(2)在12:00左右出现光合作用强度“低谷”,原因是此时________ 。
(3)如果该植物处于密闭的空间中,CO2吸收速率也如图所示,则密闭空间中CO2在________ 点浓度最高,O2在________ 点浓度最高。
(4)研究还发现,当土壤干旱时,根细胞会迅速合成某种化学物质X。有人推测根部合成X运输到叶片,能调节气孔的开闭。他们做了如下实验:从同一植株上剪取大小和生理状态一致的3片叶,分别将叶柄下部浸在不同浓度X的培养液中。一段时间后,测得的有关数据如表所示。(注:气孔导度越大,气孔开启程度越大)
①以上方案有2个不完善的地方,请指出来并加以修正________ 。
②若表中数据为方案完善后得到的结果,那么可推测,随着培养液中X的浓度增大,叶片气孔开启程度________ 。
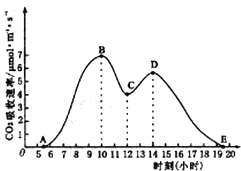
(1)在A点时植物细胞中会产生ATP的场所是
(2)在12:00左右出现光合作用强度“低谷”,原因是此时
(3)如果该植物处于密闭的空间中,CO2吸收速率也如图所示,则密闭空间中CO2在
(4)研究还发现,当土壤干旱时,根细胞会迅速合成某种化学物质X。有人推测根部合成X运输到叶片,能调节气孔的开闭。他们做了如下实验:从同一植株上剪取大小和生理状态一致的3片叶,分别将叶柄下部浸在不同浓度X的培养液中。一段时间后,测得的有关数据如表所示。(注:气孔导度越大,气孔开启程度越大)
 | 培养液中X的浓度/mol•m﹣3 | ||
| 5×10﹣5 | 5×10﹣4 | 5×10﹣3 | |
| 叶片中X的浓度/mol•g﹣1(鲜重) | 2.47 | 2.97 | 9.28 |
| 叶片中的气孔导度/mol•m﹣2•a﹣1 | 0.54 | 0.43 | 0.27 |
①以上方案有2个不完善的地方,请指出来并加以修正
②若表中数据为方案完善后得到的结果,那么可推测,随着培养液中X的浓度增大,叶片气孔开启程度
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐3】某实验小组为探究不同遮阴条件及施肥处理对油丹幼苗光饱和点(光合速率达到最大时的光照强度)和光补偿点(净光合速率为零时的光照强度)的影响,选用生长状况等相同的油丹幼苗设置了1层遮阳网遮阴组(W1)、2层遮阳网遮阴组(W2)1层遮阳网遮阴+施肥组(W3)和全光照未施肥对照组(CK),一段时间后测得各组油丹幼苗的光合特性如下表所示。回答下列问题:
(1)能反映油丹幼苗利用弱光能力的指标是_____ (填“光饱和点”或“光补偿点”),在该光照强度下,油丹幼苗的叶肉细胞中能合成ATP的场所有_____ 。
(2)某光照强度下,油丹幼苗在1层遮阳网遮阴条件下,净光合速率为零。若在此条件下增加1层遮阳网遮阴,则油丹幼苗有机物的干重会_____ 。结合以上信息,判断理由是_____ 。
(3)分析表格数据可知,施肥会_____ (填“提高”或“降低”)油丹幼苗对光能的利用率。该实验小组进一步研究发现,遮阴条件下施肥能促进幼苗中叶绿素的合成,从而影响油丹幼苗对光能的利用。从以上4组幼苗中选择合适的素材,设计实验验证该结论。
实验思路:_____ 。
预期结果:_____ 。
(4)油丹果实成熟到一定程度,其呼吸作用突然增强,然后又突然减弱,这标志着油丹果实进入衰老阶段。下列叙述错误的是_____ 。
A.油丹果实呼吸作用减弱,产生的CO2量减少
B.油丹果实的呼吸作用增强,会加快果实内糖的消耗
C.在低温条件下储藏,可降低油丹果实的呼吸作用
D.喷洒适宜浓度的乙烯利溶液,可延缓油丹果实的衰老
| 处理 | 光饱和点/(μmol·m-2·s-1) | 光补偿点/(μmol·m-2·s-1) |
| W1 | 783.75 | 11.91 |
| W2 | 947.51 | 8.96 |
| W3 | 1622.67 | 11.92 |
| CK | 1689.25 | 14.23 |
(2)某光照强度下,油丹幼苗在1层遮阳网遮阴条件下,净光合速率为零。若在此条件下增加1层遮阳网遮阴,则油丹幼苗有机物的干重会
(3)分析表格数据可知,施肥会
实验思路:
预期结果:
(4)油丹果实成熟到一定程度,其呼吸作用突然增强,然后又突然减弱,这标志着油丹果实进入衰老阶段。下列叙述错误的是
A.油丹果实呼吸作用减弱,产生的CO2量减少
B.油丹果实的呼吸作用增强,会加快果实内糖的消耗
C.在低温条件下储藏,可降低油丹果实的呼吸作用
D.喷洒适宜浓度的乙烯利溶液,可延缓油丹果实的衰老
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐1】下图为晴朗夏季一天中某植物吸收或释放CO2速率的变化曲线,f点光合午休的出现说明气孔的开闭是影响光合作用的主要因素。而气孔开闭直接与保卫细胞的水势有关,保卫细胞的水势降低. 细胞吸水使气孔张开,反之气孔关闭。
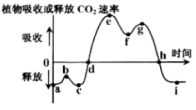
(1)水势与渗透压呈___________ 相关。
(2)结合题中所给信息分析干旱导致植物光合作用强度下降的主要原因是___________________ 。因此在大田种植中,可通过___________ 措施来缓解光合午休现象。
(3)若增加环境中的CO2浓度,f点将___________ (填“向上移”或“向下移”);h点将向___________ 移动。
(1)水势与渗透压呈
(2)结合题中所给信息分析干旱导致植物光合作用强度下降的主要原因是
(3)若增加环境中的CO2浓度,f点将
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐2】图甲表示在不同温度条件下CO2浓度对某植物净光合速率的影响;图乙表示将该种植物叶片置于适宜的光照和温度条件下,叶肉细胞中C5的相对含量随细胞间隙CO2浓度的变化曲线。请回答下列有关问题:
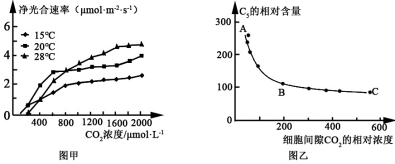
(1)据甲图可知,当CO2浓度为600μmol·L-1时,该植物叶肉细胞中能产生ATP的细胞器有______ ,当CO2浓度为200μmol·L-1、温度28℃条件下,该植物净光合速率为零,则该植物叶肉细胞中光合作用强度______ 呼吸作用强度(填“>”、“=”或“<”),在该CO2浓度时,20℃和15℃条件下该植物净光合速率明显大于28℃,原因可能是______ 。
(2)CO2在RuBP羧化酶作用下与C5结合生成C3,据此推测,RuBP羧化酶分布在______ 中。图乙中,A→B的变化是由于叶肉细胞吸收CO2速率______ (填“增加”或“减少”),B→C保持稳定的内因是受到______ 限制。
(3)研究发现,绿色植物中RuBP羧化酶具有双重活性,催化如下图所示的两个方向的反应,反应的相对速度取决于O2和CO2的相对浓度。
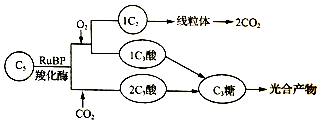
在叶绿体中,在RuBP羧化酶催化下C5与______ 反应,形成的______ 进入线粒体放出CO2,称之为光呼吸。据图推测,CO2浓度倍增可以使光合产物的积累增加,原因是______ 。
(1)据甲图可知,当CO2浓度为600μmol·L-1时,该植物叶肉细胞中能产生ATP的细胞器有
(2)CO2在RuBP羧化酶作用下与C5结合生成C3,据此推测,RuBP羧化酶分布在
(3)研究发现,绿色植物中RuBP羧化酶具有双重活性,催化如下图所示的两个方向的反应,反应的相对速度取决于O2和CO2的相对浓度。
在叶绿体中,在RuBP羧化酶催化下C5与
您最近一年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐3】科研人员研究了温度对甲植物代谢的影响,结果如图所示,图中A点表示两线的交点。回答下列问题:
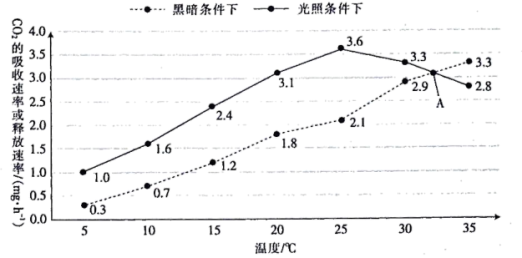
(1)黑暗条件下,甲植物的叶肉细胞中进行物质和能量转化的细胞结构是____________ ,伴随着物质变化发生的能量转化是_______________ 。
(2)据图分析,在光照条件下,A点之前,随着温度的升高,甲植物积累有机物的速率变化情况是_____________ ;A点时,甲植物制造有机物的速率____________ (填“大于”、“等于”或“小于”)其消耗有机物的速率。
(3)若要研究光照强度对甲植物光合速率的影响,则应将温度控制在____________ ℃,此时实验的检测指标除了图中所示的外,还可以是_____________ (答出一种)。
(1)黑暗条件下,甲植物的叶肉细胞中进行物质和能量转化的细胞结构是
(2)据图分析,在光照条件下,A点之前,随着温度的升高,甲植物积累有机物的速率变化情况是
(3)若要研究光照强度对甲植物光合速率的影响,则应将温度控制在
您最近一年使用:0次


