合理施用镁肥能明显提高作物的产量、改善品质。科研人员研究了镁对苗期油菜不同部位叶片生理生化指标的影响,结果如下表。
回答以下问题
(1)Rubisco酶在光合作用过程中能催化二氧化碳的固定过程,其主要存在于叶肉细胞的____ 中;植物缺镁时,叶肉细胞中五碳化合物的消耗量会____ 。
(2)据表可知,缺镁会影响光合作用的____ 阶段,判断依据是____ 。
(3)镁肥的施用量也不是越多越好,一次施用镁肥过多甚至可能导致作物死亡,这是因为____ 。
生理生化指标 | 上层叶片(新叶) | 下层叶片(老叶) | ||
镁缺乏 | 镁适宜 | 镁缺乏 | 镁适宜 | |
叶绿素含量(mg·g-1) | 0.21 | 1.12 | 0.14 | 0.67 |
Rubisco酶活性(U·g-1) | 228 | 908 | 99 | 298 |
净光合速率(μmol·m-2·s-1) | 12.2 | 16.4 | 6.8 | 16.6 |
(1)Rubisco酶在光合作用过程中能催化二氧化碳的固定过程,其主要存在于叶肉细胞的
(2)据表可知,缺镁会影响光合作用的
(3)镁肥的施用量也不是越多越好,一次施用镁肥过多甚至可能导致作物死亡,这是因为
更新时间:2024-04-24 17:28:23
|
相似题推荐
非选择题-解答题
|
适中
(0.65)
名校
【推荐1】下图1为渗透装置的示意图。图2为猪的红细胞在不同浓度的NaCl溶液中,红细胞体积和初始体积之比的变化曲线(O点对应的浓度为红细胞吸水涨破时NaCl浓度)。已知细胞膜上跨膜蛋白的合成类似于分泌蛋白,在细胞膜的跨膜蛋白中,有一种与水的跨膜运输有关的水通道蛋白,可提高水分子的运输效率。请据图回答下列问题:
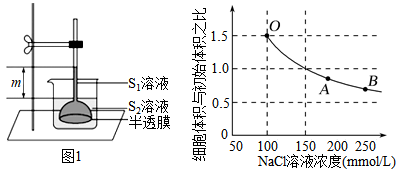
(1)图1渗透装置达到渗透平衡时S1溶液的渗透压_______________ (填“大于”“等于”或“小于”)S2溶液的渗透压。若达到平衡后吸出漏斗中高出烧杯液面的溶液,再次平衡时m将___________ (填“变大”“不变”或“变小”)
(2)根据图2可知,猪的红细胞在浓度约为_________________ 的NaCl溶液中能保持正常状态。将相同的猪的红细胞甲、乙分别放置在A点和B点对应浓度的NaCl溶液中,一段时间后红细胞乙的吸水能力________________ (填“大于”“等于”或“小于”)红细胞甲。
(1)图1渗透装置达到渗透平衡时S1溶液的渗透压
(2)根据图2可知,猪的红细胞在浓度约为
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐2】下图甲、乙、丙是不同细胞的亚显微结构模式图。请据图回答问题:(“[ ]”里填标号,“________”上填名称)·
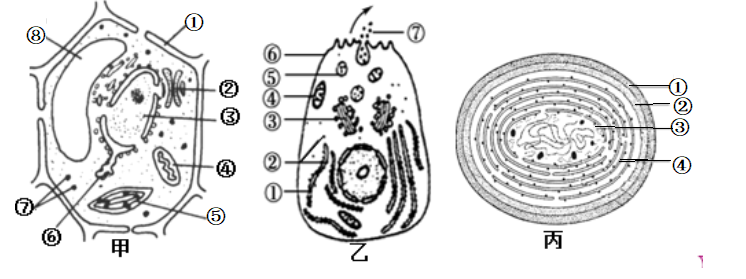
(1)甲细胞中的结构①的组成成分是________ 。如果将甲细胞放入30%的蔗糖溶液中,将会出现________ 现象。原生质层是________________________________ 。
(2)若甲细胞中含有中心体,则为________ 细胞。
(3)乙细胞中,属于生物膜系统的结构包括________ (填图中序号)。如果⑦是该细胞分泌的一种蛋白质,则参与⑦形成的细胞器有________ (填图中序号)。
(4)若用某种毒素处理乙细胞后,发现其有氧呼吸减弱,则该毒素最可损伤了[ ]________ 。
(5)若丙图中③表示的某种细胞器,则对应于甲图中[ ]________ 。
(6)为研究细胞内各种细胞器的结构和功能,需将细胞器分离,分离各种细胞器常用的方法是_________________________________ 。
(1)甲细胞中的结构①的组成成分是
(2)若甲细胞中含有中心体,则为
(3)乙细胞中,属于生物膜系统的结构包括
(4)若用某种毒素处理乙细胞后,发现其有氧呼吸减弱,则该毒素最可损伤了[ ]
(5)若丙图中③表示的某种细胞器,则对应于甲图中[ ]
(6)为研究细胞内各种细胞器的结构和功能,需将细胞器分离,分离各种细胞器常用的方法是
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐3】图甲是发生质壁分离的植物细胞图象,图乙是显微镜下观察到的某一时刻的洋葱鳞片叶的外表皮细胞,图丙表示某细胞膜结构,图中A、B、C、D、E、F表示某些物质,a、b、c、d表示物质跨膜运输方式,请据图回答下列问题:
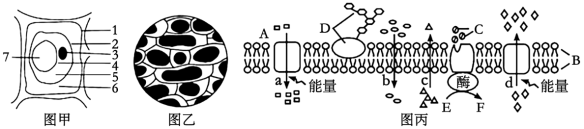
(1)将有活性的洋葱表皮细胞放入大于细胞液浓度的KNO3溶液中(此浓度不会造成细胞死亡),细胞发生质壁分离复原现象,影响该细胞离子吸收速率的因素有______________ (写出两条即可得分)。
(2)图乙此时细胞液浓度m与外界溶液浓度n的关系是___________ 。
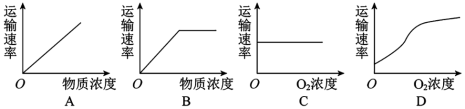
(3)图丙a、b、c、d过程主要体现细胞膜的___________ 功能。若图丙是小肠上皮细胞的细胞膜,该细胞吸收葡萄糖的跨膜运输方式可用图丙中___________ (填写a或b或c或d)表示。若图丙为胃黏膜上皮细胞的细胞膜,人在饮用雪碧(内含葡萄糖)时,下列曲线与葡萄糖跨膜运输方式相符合的是_____________ 。
(1)将有活性的洋葱表皮细胞放入大于细胞液浓度的KNO3溶液中(此浓度不会造成细胞死亡),细胞发生质壁分离复原现象,影响该细胞离子吸收速率的因素有
(2)图乙此时细胞液浓度m与外界溶液浓度n的关系是
(3)图丙a、b、c、d过程主要体现细胞膜的
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐1】铜绿微囊藻是淡水中常见的一种蓝藻,它们能够将大气中的氮固定为可利用的氮源。当其在富营养化的水体中大量繁殖时,会导致水体变质,这种现象俗称“水华”。研究者尝试用中草药黄连提取物——小檗碱来抑制铜绿微囊藻的生长。
(1)在铜绿微囊藻培养液中添加 4mg/L 小檗碱,分别在黑暗(图甲)和正常光照(图乙)两种条件下培养,每 24h 取样测定培养液中藻细胞密度,结果如下图所示。
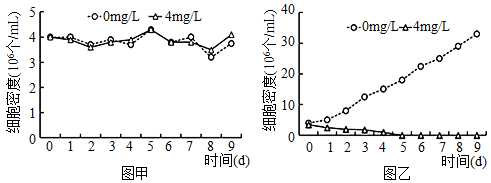
根据实验结果,从生理过程分析,可推断出___________________________________________ (结论)。
(2)已知光合作用的光反应主要在光系统Ⅱ(PSⅡ)和光系统Ⅰ(PSⅠ)内进行。这两个光系统协同完成受光激发推动的高能电子从H2O向NADP+的传递,最终形成的__________ 用于暗反应中__________ 过程。为进一步研究小檗碱的作用机制,测定了铜绿微囊藻的全链电子传递活性、PSII 和 PSI 电子传递活性,结果见下表。
结果表明,小檗碱抑制铜绿微囊藻光合作用的主要作用位点位于______________ 。
(3)另有实验表明,小檗碱无法对绿藻类的光合作用发挥效应。从细胞结构的角度分析,小檗碱对绿藻类无效的原因可能是___________________________ 。
(4)为有效控制铜绿微囊藻的增长,一方面需要对污水中含______ (填“氮”或“磷”)量进行控制,另一方面可以在排污口附近种植芦苇、莲等挺水植物,这体现了生物多样性的_______ 价值。
(1)在铜绿微囊藻培养液中添加 4mg/L 小檗碱,分别在黑暗(图甲)和正常光照(图乙)两种条件下培养,每 24h 取样测定培养液中藻细胞密度,结果如下图所示。
根据实验结果,从生理过程分析,可推断出
(2)已知光合作用的光反应主要在光系统Ⅱ(PSⅡ)和光系统Ⅰ(PSⅠ)内进行。这两个光系统协同完成受光激发推动的高能电子从H2O向NADP+的传递,最终形成的
小檗碱浓度 (mg/L) | 光合电子传递活性(µmolO2·mgChla-1·h-1) | ||
全链 | PSⅡ | PSⅠ | |
0 | 132.63 | 248.06 | 62.75 |
0.5 | 108.53 | 103.85 | 69.23 |
结果表明,小檗碱抑制铜绿微囊藻光合作用的主要作用位点位于
(3)另有实验表明,小檗碱无法对绿藻类的光合作用发挥效应。从细胞结构的角度分析,小檗碱对绿藻类无效的原因可能是
(4)为有效控制铜绿微囊藻的增长,一方面需要对污水中含
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐2】为研究某种植物对盐的耐受性,科学家测定了不同盐浓度对该植物的净光合速率和光合色素含量的影响,结果如下表所示。回答问题:
(1)从某植物叶片中提取光和色素时,加入CaCO3的目的是______________ ;与光合作用有关的色素分布在叶绿体的_______________________ 上。
(2)光合作用的过程可分为光反应和暗反应两个阶段,其中光反应为暗反应提供__________ 。
(3)据表分析,与低盐相比,当NaCl浓度在200~250mmol/L时,净光合速率________ 。
原因是:
①__________________________ ,造成光反应减弱。
②根细胞周围盐浓度增高,根细胞会因渗透作用失水,导致叶片萎蔫,气孔关闭,使_______________ ,造成暗反应受阻。
| NaCl浓度(mmol/L) | 净光合速率(μmolCO2·m-2·s-1) | 光合色素含量(mg/g) | |
| 叶绿体 | 类胡萝卜素 | ||
| 0 | 6.0 | 2.3 | 0.75 |
| 25 | 6.5 | 2.6 | 1.00 |
| 50 | 7.0 | 2.8 | 0.60 |
| 100 | 5.9 | 2.4 | 0.55 |
| 150 | 5.7 | 2.0 | 0.50 |
| 200 | 2.5 | 1.8 | 0.48 |
| 250 | 2.0 | 1.5 | 0.45 |
(1)从某植物叶片中提取光和色素时,加入CaCO3的目的是
(2)光合作用的过程可分为光反应和暗反应两个阶段,其中光反应为暗反应提供
(3)据表分析,与低盐相比,当NaCl浓度在200~250mmol/L时,净光合速率
原因是:
①
②根细胞周围盐浓度增高,根细胞会因渗透作用失水,导致叶片萎蔫,气孔关闭,使
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐3】科研人员对温室栽种的番茄进行了相关研究,在不同条件下测定了其光合速率(用CO2的吸收速率表示),变化曲线如下图所示。请回答下列问题:
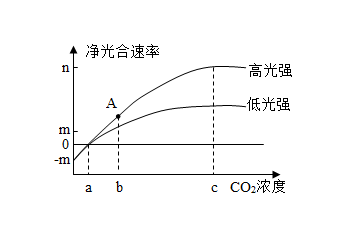
(1)叶肉细胞中叶绿体中的色素能够捕获光能,这些能量经过转换,最终转化为__________________________ 。CO2浓度小于a时,图示的两种光强下番茄植株呼吸作用产生的CO2量____________ (填“大于”“等于”或“小于”)光合作用固定的CO2量。CO2浓度为b时限制番茄光合速率的环境因素主要是____________________ 。
(2)据图分析,番茄在高光强条件下,CO2浓度为c和b时光合速率差值较大,而弱光强条件下这种差值明显缩小,试分析原因:______________________________ 。
(3)某研究小组发现适当遮光有利于番茄幼苗的生长,对此现象他们给出了一个解释,适当遮光可以使番茄幼苗的叶绿素含量增加。请参照色素的提取和分离原理设计一个简单的实验,来探究他们的解释是否正确,简要写出实验思路和预期实验结果。
实验思路:_____________________________ 。
预期实验结果:___________________________ 。
(1)叶肉细胞中叶绿体中的色素能够捕获光能,这些能量经过转换,最终转化为
(2)据图分析,番茄在高光强条件下,CO2浓度为c和b时光合速率差值较大,而弱光强条件下这种差值明显缩小,试分析原因:
(3)某研究小组发现适当遮光有利于番茄幼苗的生长,对此现象他们给出了一个解释,适当遮光可以使番茄幼苗的叶绿素含量增加。请参照色素的提取和分离原理设计一个简单的实验,来探究他们的解释是否正确,简要写出实验思路和预期实验结果。
实验思路:
预期实验结果:
您最近半年使用:0次
非选择题-解答题组
|
适中
(0.65)
名校
【推荐1】薄荷是大家熟知的一种多年生草本植物,可食用,可做药材,有清热明目等功效。薄荷喜阳喜温,在荫蔽寒冷环境中生长受抑制。科研小组利用薄荷开展了相关实验。
Ⅰ.通过遮光实验测定薄荷有关生理指标如下表:
(1)该实验为________ (填“对照实验”或“对比实验”)。实验结果可知,荫蔽条件下叶肉细胞中的_____ 含量明显增加,推测这一变化有利于叶片吸收和利用光能,促进光反应阶段的能量转换。
(2)与自然光下的结果相比较,薄荷叶片在荫蔽条件下真(总)光合速率的变化是________ (填“增大”“减小”或“几乎不变”),据表中数据分析,其原因是________ 。
Ⅱ.科研小组利用低温处理薄荷,研究低温胁迫对薄荷光合特性的影响(12℃为对照“CK”,3个低温处理组“L1、L2、L3”,分别为-2℃、-4℃、-6℃),结果如下表所示:
(3)低温下薄荷叶片出现缺绿现象,综合表中Mg2+含量结果分析,低温下薄荷叶片表现出缺绿的主要原因是________ 。
(4)据表分析可知,3个实验组的净光合速率比CK组低。原因是否都是低温降低了光合作用有关酶的活性?________ (填“是”或“否”)。请作出解释:________
(5)研究发现,植物体中OST1蛋白能够促进气孔打开。科研人员获得了光合速率明显降低的OST1基因功能缺失的突变体植株,并推测OST1基因表达受到光合作用产物(如蔗糖等)的调控,进而影响气孔的开放程度。请以野生型和突变体作物作为实验材料,设计实验加以验证(补全下列实验思路即可):
________
一段时间后检测叶片中气孔的开放程度。
Ⅰ.通过遮光实验测定薄荷有关生理指标如下表:
实验条件 | 叶绿素a含量(mg·g-1) | 叶绿素b含量(mg·g-1) | 呼吸速率(μmolCO2·m-2·s-1) | 净光合速率(μmolCO2·m-2·s-1) |
荫蔽 | 1.65 | 0.72 | 2.11 | 20.01 |
自然光 | 1.27 | 0.70 | 3.46 | 18.69 |
(2)与自然光下的结果相比较,薄荷叶片在荫蔽条件下真(总)光合速率的变化是
Ⅱ.科研小组利用低温处理薄荷,研究低温胁迫对薄荷光合特性的影响(12℃为对照“CK”,3个低温处理组“L1、L2、L3”,分别为-2℃、-4℃、-6℃),结果如下表所示:
处理 | 叶片中Mg2+含量(ug·g-1·FW) | 茎中Mg²⁺含量(ug· g-1·FW) | 净光合速率(μmol·m-2·s) | 气孔导度(μmmol·m-2. S-1) | 蒸腾速率(μmol·m-2● s-1) | 胞间CO2浓度(μ mol·mol·) |
CK | 580 | 16 | 16.2 | 512.6 | 3.7 | 261.4 |
L₁ | 520 | 215 | 13.3 | 376.1 | 2.6 | 230.6 |
L₂ | 506 | 242 | 8.6 | 272.8 | 1.8 | 276.3 |
L₃ | 482 | 273 | 6.5 | 183.6 | 1.6 | 310.5 |
(4)据表分析可知,3个实验组的净光合速率比CK组低。原因是否都是低温降低了光合作用有关酶的活性?
(5)研究发现,植物体中OST1蛋白能够促进气孔打开。科研人员获得了光合速率明显降低的OST1基因功能缺失的突变体植株,并推测OST1基因表达受到光合作用产物(如蔗糖等)的调控,进而影响气孔的开放程度。请以野生型和突变体作物作为实验材料,设计实验加以验证(补全下列实验思路即可):
一段时间后检测叶片中气孔的开放程度。
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
名校
【推荐2】如图表示在不同温度下,测定某植物叶片重量变化情况(均考虑为有机物的重量变化)的操作流程及结果,据图分析回答问题:
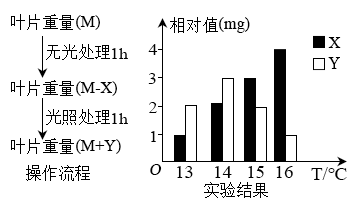
(1)从图分析可知,该植物的呼吸速率可表示为______ ,实际光合速率可表示为______ 。在13~16 ℃之间,随着温度的升高,呼吸作用强度___________ (增强、减弱、不变),实际光合作用的强度__________ (增强、减弱、不变)。
(2)恒定在上述____ ℃温度下,维持10小时光照,10小时黑暗,该植物叶片增重最多。
(1)从图分析可知,该植物的呼吸速率可表示为
(2)恒定在上述
您最近半年使用:0次
非选择题-解答题
|
适中
(0.65)
【推荐3】当其他条件均最适宜时,北沙参在不同温度和光照强度条件下的光合速率如图所示,光合速率以干重增加速率(mg/单位叶面积•h)表示.请分析回答下列问题:
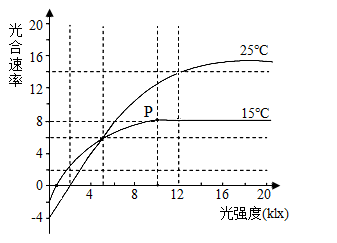
(1)图中表明影响植物光合作用强度的环境因素是________ ,25℃条件下,5klx光照强度时的真正光合速率________ (选填“大于”“等于”或“小于”)P点代表的真正光合速率
(2)15℃条件下,P点以后北沙参光合速率不再继续增大的限制因素是________ ;若将CO2浓度降低为大气中CO2浓度值,则叶绿体中[H]合成速率将会________ (选填“变大”“不变”或“变小”),P点向________ 方移动
(1)图中表明影响植物光合作用强度的环境因素是
(2)15℃条件下,P点以后北沙参光合速率不再继续增大的限制因素是
您最近半年使用:0次


