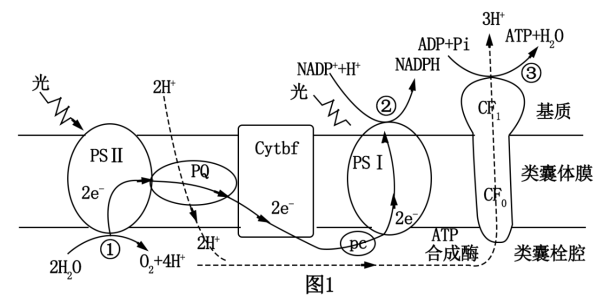
(2)春美叶片中的色素利用光能,将水分解成
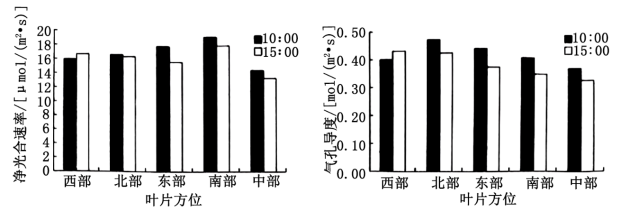
(3)10:00和15:00时光照强度相当,春美的呼吸速率基本不变。除西部叶片外,在10:00叶片的净光合速率均
(4)中部叶片净光合速率最低,推测可能的原因是
品种 | 地点 | 净光合速率(μmol·m-2·s-1) | 气孔导度/(mol·m-2·s-1) | 胞间CO2浓度(μL·L-1) | 果实成熟期(月/日) |
布鲁克斯 | 临海 | 2.92 | 0.0224 | 158.26 | 5/3~5/10 |
仙居 | 9.35 | 0.0744 | 167.28 | 5/10~5/20 | |
黄岩 | 13.73 | 0.1477 | 206.22 | 5/22~5/26 | |
拉宾斯 | 临海 | 4.66 | 0.0399 | 193.07 | 4/30~5/9 |
仙居 | 8.44 | 0.0664 | 164.43 | 5/7~5/13 | |
黄岩 | 13.69 | 0.1676 | 229.56 | 5/14~5/22 | |
红蜜 | 临海 | 5.88 | 0.0600 | 197.54 | 5/6~5/12 |
仙居 | 11.61 | 0.1090 | 197.54 | 5/10~5/14 | |
黄岩 | 16.88 | 0.2610 | 254.49 | 5/21~5/31 |
(2)光照强度主要影响光合作用的
(3)由上表可知,在不同海拔高度条件下,甜樱桃光合特性存在显著差异,海拔越高,相对来看净光合速率越

(2)图1中灯与小球藻培养液之间盛水玻璃柱的作用是
(3)通过纸层析法分离混合代谢产物的原理是
(4)结果发现:照光30秒,14C分布于许多化合物中,缩短到1秒时,几乎所有的14C都集中在3-磷酸甘油酸上,这说明
(5)光通过光反应改变叶绿体的内部环境,间接地影响酶的活性。例如,光促进H+从叶绿体基质进入类囊体腔内,同时交换出Mg2+,使叶绿体基质中的H+和Mg2+浓度正适合与暗反应相关酶的活性。在弱光环境下,植物暗反应速率降低的原因是
光源 | 叶绿素含量(mg·g-1) | 气孔导度(mmol·m-2·s-1) | 净光合速率(μmol·m-2·s-1) | 单果质量(g·果-1) | 结果数(个·株-1) |
CK组 | 1.13 | 308 | 16.04 | 12.11 | 9.9 |
T1组 | 2.31 | 313 | 16.32 | 14.51 | 13.2 |
T2组 | 1.79 | 310 | 15.39 | 18.07 | 12.7 |
(1)叶绿素含量升高后会通过增加
(2)假设实验过程中各组实验材料呼吸作用强度不变且相同,与CK组相比,T1组的净光合速率升高,除该组植株叶片中叶绿素含量升高、固定光能增多外,其他可能原因有
(3)从光合色素的吸收光谱考虑,科研人员建议增设T3组,预期T3组的处理方法为
(4)上述三组实验结果显示,在种植同样株数的草莓时,产量最高的一组是
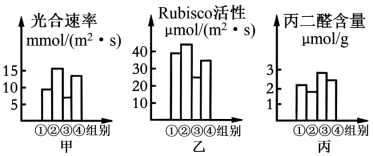
(1)Rubisco催化CO₂固定的反应产物是
(2)增产胺对Rubisco活性的影响在遮光条件下更大,判断依据是
(3)为研究遮光和不遮光条件下增产胺对甜瓜叶绿素含量的影响,如何继续进行实验,实验思路是:
(2)强光条件下,过剩的光能会对PSⅡ复合体造成损伤,导致光合作用强度减弱。遭受强光损伤的拟南芥幼叶细胞中,叶绿素酶(CLH)基因表达量明显上升,科研人员为研究幼叶应对强光影响的机制,分别测定野生型(WT)、CLH基因缺失的突变型(clh-1)和CLH基因过量表达的突变型(clh-2)拟南芥在强光照射后的生存率,结果如图2所示。据图可知,CLH基因可以
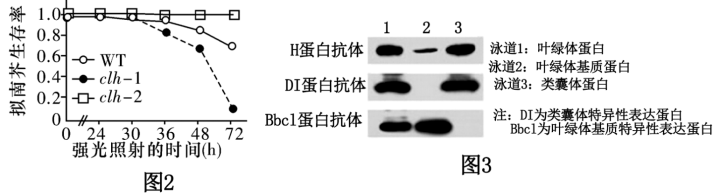
②科研人员用特定抗体检测H蛋白在叶绿体内的分布,结果如图3所示(各泳道的蛋白上样量均保持一致),依据实验结果可以得出的结论是
③H蛋白是一种热应激蛋白(温度升高时表达),调控叶绿体基因编码的RNA聚合酶的活性。据此推测,H基因突变体在30℃时叶子呈白色的原因是
项目 | 甲组 | 乙组 | 丙组 |
库源比 | 1/2 | 1/4 | 1/6 |
净光合速率/(μmol·m-2·s-1) | 9.31 | 8.99 | 8.75 |
果实中含13C光合产物/mg | 21.96 | 37.38 | 66.06 |
单果重/g | 11.81 | 12.21 | 19.59 |
(2)光反应产生的
(3)已知卡尔文循环的中间产物三碳糖磷酸既可用于合成淀粉,还可经叶绿体膜上的磷酸转运器运至细胞质基质中参与蔗糖的合成。光合产物从“源”向“库”运输的物质形式主要是
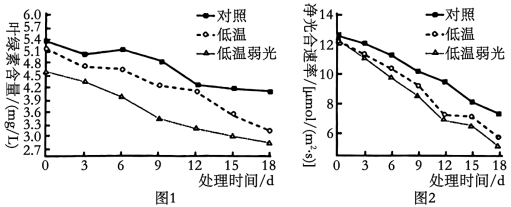
(2)由图1、2可知,相对于低温处理,在低温弱光处理下,净光合速率下降得更明显,主要是因为
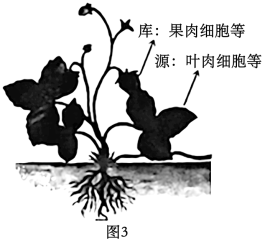
(3)如图3所示,在植物体内,制造并输出有机物的组织器官被称为“源”,接纳有机物用于生长或贮藏的组织器官被称为“库”。若要验证“源”、“库”之间存在上述物质代谢关系,可以使用14CO2饲喂草莓叶片,检测
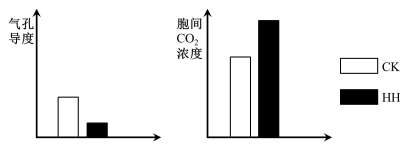
(1)高温、强光条件下会使叶绿素分解,为验证这一现象,取高温、强光条件和适宜条件下的等量该植物叶片,用
(2)实验测得高温、强光条件下植物光合速率低于适宜条件,据图分析引起两组光合速率差异的主要因素
(3)若高温、强光条件下植物细胞会产生自由基破坏生物膜结构,据此分析HH组恢复适宜条件后短期内光合速率仍然较低,其原因可能是
光合特征参数 | 丽薯6号 | 中薯20 | ||
自然光照 | 遮光70% | 自然光照 | 遮光70% | |
净光合速率/(μmol·m-2·s-1) | 21.45 | 12.26 | 24.20 | 17.52 |
气孔导度/(mol·m-2·s-1) | 0.40 | 0.18 | 0.57 | 0.45 |
光补偿点/(μmo·m-2·s-1) | 45.83 | 19.05 | 27.31 | 17.75 |
呼吸速率/(μmol·m-2·s-1) | 2.64 | 1.30 | 1.47 | 1.11 |
(1)遮光会使两个品种马铃薯的最大光合速率降低,从光反应的角度分析,具体原因可能是
(2)若中薯20处于光补偿点时,将丽薯6号置于其相同条件下能否生长?
(3)以上两个品种中,更适合与玉米等作物间作种植的是


