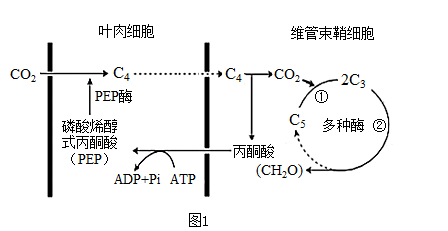
(1)实验室研究玉米叶片中色素种类及相对含量。提取色素时,为防止叶绿素被破坏,可以加入
(2)若玉米的维管束鞘细胞叶绿体中只能进行碳反应,推测其可能缺少的结构是
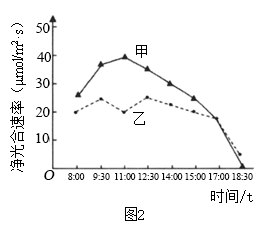
(3)上午9:00时,突然降低环境中CO2浓度的一小段时间内,图2中玉米细胞和小麦细胞中C3含量的变化分别是
(4)综上分析,玉米植株净光合速率为图2中曲线
相似题推荐
组别 | 叶绿素/(mg·g-1) | 类胡萝卜素/(mg·g-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/(μmol·mol-1) | 平均每株产量/(kg/株) |
正常土壤 | 0.59 | 0.11 | 8.90 | 386.67 | 301.33 | 12 |
酸性土壤 | 0.40 | 0.07 | 6.20 | 327.33 | 350.67 | 9 |
酸性土壤+硝酸铵钙 | 0.58 | 0.12 | 8.69 | 386.63 | 302.12 | 13 |
(1)本实验的自变量是
(2)与正常土壤组相比,酸性土壤组导致黄瓜净光合速率显著下降
(3)在提取到黄瓜叶片光合色素后,可通过对比各组黄瓜叶片对某种颜色的光的吸收率来计算叶片中的叶绿素含量。为减少其他光合色素的干扰,“某种颜色的光”最可能是
(4)与生长在正常土壤中的黄瓜植株相比,生长在酸性土壤+硝酸铵钙土壤中的黄瓜植株单位时间内有机物的制造量
(1)小麦叶肉细胞中的光合色素主要有
(2)在小麦叶肉细胞中存在着下图1所示的代谢过程:

①现代细胞分子生物学研究发现RuBP羧化/加氧酶由8个大亚基(L)和8个小亚基(S)组成。高等植物细胞中L由叶绿体基因编码并在叶绿体中合成,S由细胞核基因编码并在
②RuBP羧化/加氧酶可催化CO2与RuBP(C5)结合,生成2分子C3,影响该反应的外部因素,除光照条件外还包括
③RuBP羧化/加氧酶还可参与催化C5与O2反应产生乙醇酸,乙醇酸中的碳又重新生成CO2和C3。该过程使细胞内O2/CO2的比值
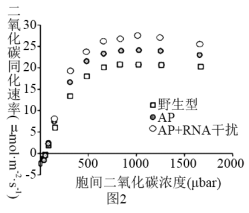
(3)根据对光呼吸机理的研究,科研人员利用基因编辑手段设计了只在叶绿体中完成的光呼吸替代途径AP(依然具有降解乙醇酸产生CO2的能力)。同时,利用RNA干扰技术,降低叶绿体膜上乙醇酸转运蛋白的表达量。检测三种不同类型植株的光合速率,实验结果如图2所示。据此回答:
当胞间CO2浓度较高时,三种类型植株中,AP+RNA干扰型光合速率的最高的原因可能是:
(1)从新鲜胡萝卜中提取胡萝卜素时,萃取前需对胡萝卜进行粉碎和
(2)用纸层析法分离叶绿体中的色素时,胡萝卜素随层析液在滤纸条上的扩散速度
(3)科研人员测定了夏季不同天气条件下胡萝卜植株光合速率的变化情况,结果如图所示,请分析回答:
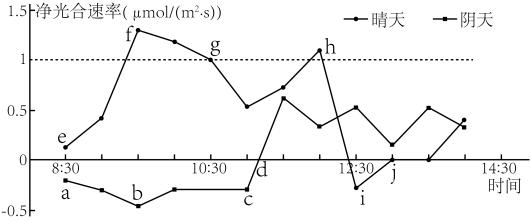
①图中j点时,叶肉细胞中叶绿体产生的氧气会扩散到
②阴天时,a~b时间段植株净光合速率变化的原因是
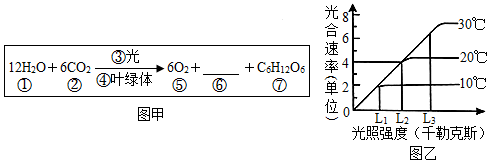
(1)蓝莓中的花青素主要存在于蓝莓细胞的
(2)参照图甲中光合作用反应式可知,影响蓝莓光合作用的因素有光照强度和温度等环境因素,除此之外,影响绿色植物光合作用的内部因素还与
(3)图乙中的光合速率是指
(4)已有研究表明,四季秋海棠在秋季低温和短日照条件下叶片会变红。若要验证“此条件下,诱导四季秋海棠叶片合成花色素苷的物质基础是 ABA 的积累”,实验组应选择的操作至少应包括
a.长日照条件下培养 b.短日照条件下培养 c.等日照条件下培养
d.低温(白天 15℃夜晚 6℃)培养 e.常温(白天 25℃夜晚 15℃)培养
f.外施 10μmol/LABA g.外施 500μmol/LABA
h.测定花色素苷含量 i.测定 ABA 含量
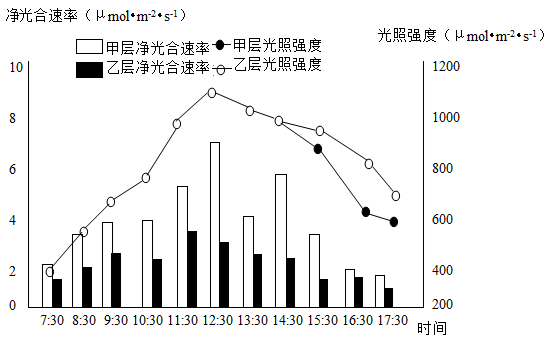
(1)植物叶肉细胞将光能转化为化学能的场所是
(2)甲表示林冠
(3)根据7:30~14:30测得的相关数据,可判断甲层叶片对光能的利用率更高,理由是
(4)每分子叶绿素含有一个Mg2+,可被H+、Cu2+等置换。在用该植物的绿叶做“色素的提取与分离”实验时,滤液用5%的HCI处理一段时间后,其颜色与研磨时未加
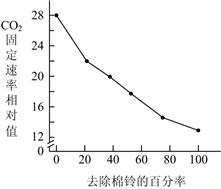

图 1 图 2
(1)光合作用暗反应为光反应提供
(2)由图1可知,随着去除棉铃百分率的提高,叶片光合速率
(3)由图2可知,去除棉铃后,植株叶片中
(4)综合上述结果可推测,叶片中光合产物的积累会
(5)一种验证上述推测的方法为:去除植株上的棉铃并对部分叶片遮光处理,使遮光叶片成为需要光合产物输入的器官,检测
处理 | 指 标 | ||||||
Pn/(μmol·m-²s-') | 叶绿素含量/(mg/g) | 类胡萝卜素含量/(mg/g) | Gs/(mmol·m-2 · s-¹) | 单果质量/(g) | 结果数/ (个株¹) | 果实成熟日期 | |
对照 | 16.04 | 1.13 | 0.08 | 307.99 | 12.11 | 9.90 | 02-05 |
T1 | 16.32 | 2.31 | 0.15 | 343.09 | 14.51 | 13.20 | 02-04 |
T2 | 17.03 | 1.93 | 0.14 | 330.91 | 18.33 | 13.60 | 01-31 |
T3 | 15.39 | 1.79 | 0.11 | 340.53 | 18.07 | 12.70 | 02-01 |
T4 | 14.06 | 1.99 | 0.14 | 333.74 | 12.30 | 13.90 | 01-31 |
(1)在光反应中,被夺去电子的物质是
(2)市面上的补光灯中经常出现红光荧光灯的原因是
(3)T1处理
(4)从产量指标分析该草莓种植园最应该选择
处理 | 净光合速率(μmol·m2·s-1) | 叶绿素含量(mg·g-1) | 气孔导度(mol·m-2·s-1) | 胞间CO2浓度(uL·m2·L-1) | RuBP羧化酶活性(nmol·min-1·g-1) |
对照 | 22.9 | 24.2 | 605 | 351 | 172 |
PEG | -0.69 | 14.7 | 34 | 505 | 78 |
PEG+FA | 11.7 | 20.3 | 108 | 267.8 | 119 |
(1)黄瓜叶片中的光合色素有
(2)据表分析,干旱条件下喷施一定浓度的FA可以提高暗反应速率的原因是
(3)在叶绿体中叶绿素水解酶与叶绿素是单独分布的。干旱会导致叶绿体大量变形、基粒片层完全解体,使光合作用速率降低,据此分析,FA可通过
(4)研究人员探究了CO2浓度对黄瓜幼苗光合速率的影响,将黄瓜幼苗分别进行不同实验处理:甲组提供大气CO2浓度(375μmol·mol-1);乙组先在CO2浓度倍增环境(750μmol·mol-1)中培养60d,然后在测定前一周恢复为大气CO2浓度,其他条件相同且适宜。在晴天上午测定各组的光合速率,结果乙组光合速率比甲组低,推测原因可能是
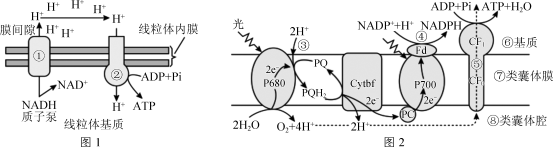
 图3
图3(1)图中①②③⑤过程都表示质子的跨膜运输,其中属于主动运输的过程是
(2)据图2判断,水的光解发生在
(3)图3叶肉细胞进行光合作用时,CO2与C5结合产生三碳酸,继而还原成三碳糖(C3),其中一部分C3运到
(4)在光照过强时,叶肉细胞必须耗散掉叶绿体吸收的过多光能,避免细胞损伤。草酸乙酸/苹果酸穿梭可有效地将光照产生的
(5)为研究线粒体对光合作用的影响,用寡霉素(电子传递链抑制剂)处理大麦,实验方法是:取培养10~14 d大麦苗,将其茎浸入添加了不同浓度寡霉素的水中,通过蒸腾作用使药物进入叶片。光照培养后测定,计算光合放氧速率(单位时间内每毫克叶绿素释放氧气的量)。请完成下表。
实验步骤的目的 | 简要操作过程 |
配制不同浓度的寡霉素丙酮溶液 | 寡霉素难溶于水,需先溶于丙酮,配制高浓度母液,并用丙酮稀释成不同药物浓度,用于加入水中 |
设置寡霉素为单一变量的对照组 | ① |
② | 对照组和各实验组均测定多个大麦叶片 |
光合放氧测定 | 用氧电极测定叶片放氧 |
③ | 称重叶片,加乙醇研磨,定容,离心,取上清液测定 |
| 光照阶段 | 黑暗阶段 | |||||||||||
| 时间 C02:浓度 (  L/L) L/L) | 8:00 | 9:00 | 10:00 | 11:00 | 12:00-24:00 | 1:00 | 2:00 | 3:00 | 4:00 | 5:00 | 6:00 | 7:00 |
| 660 | 420 | 300 | 180 | 180 | 270 | 360 | 450 | 520 | 570 | 610 | 640 | |
(1)光照培养阶段CO2浓度下降的生理原因是
(2)11:00-24:00时间段,幼苗叶肉细胞中能产生ATP的结构包括
(3)整个黑暗培养阶段幼苗呼吸速率的变化情况是
(4)若给该密闭容器中的植物提供
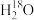 ,一段时间后楦物幼苗内出现了
,一段时间后楦物幼苗内出现了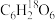 ,请用分子式、箭头、文字简要表示该过程(文字以表明生理活动):
,请用分子式、箭头、文字简要表示该过程(文字以表明生理活动):
(1)在一定条件下,若用甲图装置测定获得图乙中的G点对应的值,则甲图中B三角瓶内应放置的液体为
 ”或“
”或“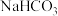 ”)溶液,测定时甲装置应置于
”)溶液,测定时甲装置应置于(2)据乙图分析,在该实验条件下,此绿色植物的光饱和点是
 浓度,图乙中的E点将向
浓度,图乙中的E点将向(3)丙图中的a、b、c、d表示物质,则a表示

(1)上图所示的生理过程表示光合作用的
(2)若将光照撤离,短时间内叶绿体中RuBP的含量


