胡杨是生长在沙漠中的乔木树种。胡杨林具有喜光耐热、耐旱涝、耐盐碱等特点,对沙漠环境具有很强的适应性,其强大的根系在土层内向四周延伸,并直达潜水面。当胡杨体内盐分积累过多时,便从树干的节疤和裂口处将盐分自动排泄出去,堪称“拔盐改土”的“土壤改良功臣”。下图示意胡杨林景观及胡杨林根质量密度(单位体积土壤中根系的质量)的垂直分布。

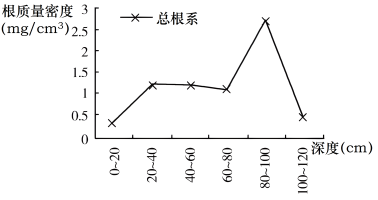
(2)指出胡杨林所属的植被类型及其具有的生态价值。
(3)胡杨幼树上的叶片细长如柳叶,老树的叶片大而厚,表面有蜡质层。结合胡杨树叶形态,说出其适应的气候特点。
山地垂直带内部的分异规律,日益为地理学研究所重视.在山地苔原带,植物多样性存在随海拔升高呈单峰变化的规律:在山地苔原带下部,少数植物种类通过种间竞争获得优势,植物多样性较低;随着海拔升高,环境压力变大,种间竞争减弱,植物多样性升高;在更高海拔区域,适宜生存的植物种类减少.地理科考队调查某山峰的苔原带(海拔2000-2600米)时发现,该苔原带部分地区存在干扰,导致优势植物数量减少,植物多样性异常;阴、阳坡降水量与坡度差别不大,但植物多样性差异显著(如图)
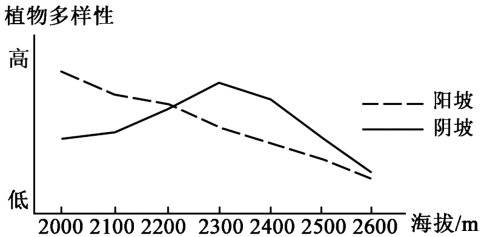
(1)确定该苔原带遭受干扰的坡向和部位,以及干扰强度的垂直分布特征.
(2)分析与阴坡相比,苔原带阳坡地表温度和湿度的特点及产生原因.
(3)说明从2300米至2600米,阴、阳破植物多样性差异逐渐缩小的原因
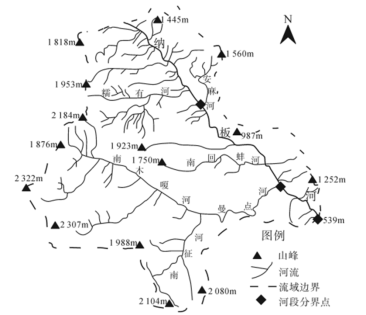
纳板河流域是云南省西双版纳族自治州境内一个典型的山区小流域,是我国第一个按小流域生物圈保护理念规划建设的自然保护区,2002年4月晋升为国家级自然保护区。区内生物多样性丰富,主要保护对象为以热带雨林为主体的森林生态系统及珍稀野生动植物。纳板河是澜沧江右岸一级支流,上、中、下游河段河道特征差异较大。图示意纳板河水系,表为纳板河各河段特征的数据。
| 河段 | 范围 | 河长/km | 落差/m | 河谷地貌类型 |
| 上游 | 纳板河源至安麻河口 | 11.37 | 960 | 岩槛,跌水,瀑布 |
| 中游 | 安麻河口至曼点河口 | 9.37 | l10 | 边滩,河漫滩,阶地 |
| 下游 | 曼点河口至纳板河口 | 3.76 | 4 | 边滩,河漫滩,阶地 |
(2)与中下游相比,说明纳板河上游的流量、流速特征及河谷地貌主要成因。
(3)分析纳板河流域生物多样性丰富的原因。
高山林线作为郁闭森林和高山植被之间的生态过渡带,是极端环境条件下树木生存的界限。色季拉山位于雅鲁藏布江大拐弯西北侧,地处藏东南湿润气候区与半湿润气候区的分界处,受印度洋季风的影响,其林线处森林植物群落类型呈现多样性特征。

(1)比较色季拉山阳坡和阴坡的物种多样性状况。
(2)说明色季拉山阴坡林线处物种丰富度的特点及形成原因。
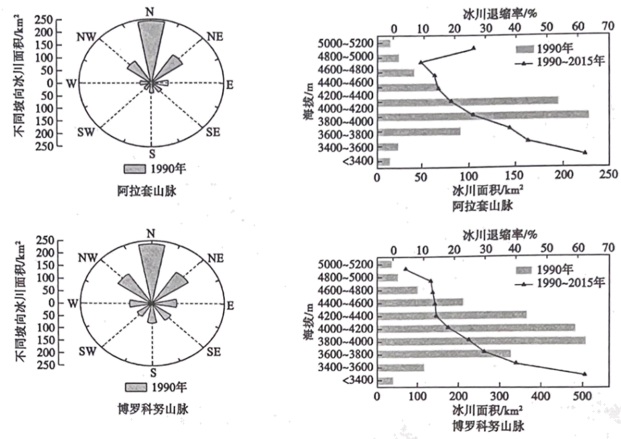
冰川面积的变化与气候、地形等因素有关,较小的冰川对气候变化更敏感。天山的阿拉套山脉和博罗科努山脉位于中国新疆维吾尔自治区,山脉均为东西延伸,两山脉形成了不同规模的冰川(下图)。1990~2015年,该区域平均气温呈上升趋势,冰川面积不断减小,研究发现阿拉套山脉比博罗科努山脉冰川退缩更快。表为这两座山脉不同规模等级的冰川面积和数量。
| 阿拉套山脉 | 博罗科努山脉 | |||||||
| 规模等级/km² | 1990年 | 2015年 | 1990年 | 2015年 | ||||
| 数量/条 | 面积/km² | 数量/条 | 面积/km² | 数量/条 | 面积/km² | 数量/条 | 面积/km² | |
| Ⅰ≤0.1 | 470 | 19.80 | 649 | 20.86 | 373 | 20.21 | 912 | 53.97 |
| Ⅱ0.1~0.5 | 442 | 109.76 | 381 | 89.78 | 933 | 259.25 | 950 | 265.96 |
| Ⅲ0.5~2 | 211 | 209.06 | 181 | 177.21 | 686 | 638.56 | 509 | 465.53 |
| Ⅳ2~5 | 43 | 136.72 | 26 | 96.55 | 135 | 417.94 | 106 | 322.69 |
| Ⅴ>5 | 21 | 236.37 | 12 | 149.05 | 74 | 889.84 | 54 | 613.20 |
| 合计 | 1187 | 711.71 | 1249 | 533.45 | 2201 | 2235.80 | 2531 | 1721.35 |
(1)据图说明这两座山脉冰川的分布特点。
(2)计算2015年与1990年博罗科努山脉V等级冰川的面积占比的差值,并简述1990~2015年该山脉不同规模等级冰川的变化特征。
(3)试分析阿拉套山脉比博罗科努山脉冰川退缩速度快的原因。
土壤容重也称土壤假比重,指一定容积的土壤(包括土粒及粒间的孔隙)烘干后的重量与同容积水重的比值,它与土壤质地、土壤水分、压实状况、土壤颗粒密度、土壤有机质含量及各种土壤管理措施有关。科研人员在野外调查祁连山中段山地表层土壤容重时发现:不同植被类型下,土壤容重不同;随着土壤深度的增加土壤容重发生变化。图1为祁连山中段山地南北坡垂直自然带谱示意,图2为祁连山中段不同植被类型0~10cm土壤容重统计。
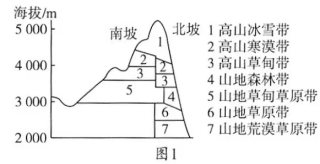
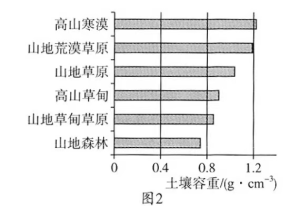
(1)指出祁连山中段山地土壤生产力较高的坡向并描述其表层土壤容重的垂直分布特征。
(2)与海拔同为3300米的山地北坡相比,指出南坡的土壤容重特点并分析其原因。
(3)说明在同一植被类型下的土壤容重随土层深度的变化情况及其原因。
非洲自然环境复杂多样,刚果盆地雨林茂密,马达加斯加岛动植物种类丰富,其中80%为特有物种。图为“非洲气候类型图及马达加斯加岛位置示意图”。
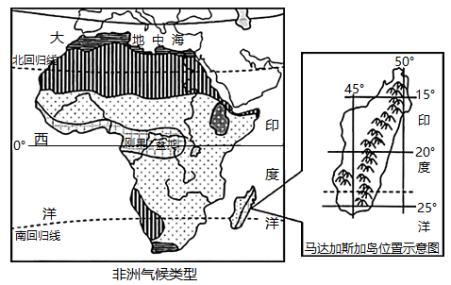
(1)根据材料推测马达加斯加岛降水的空间分布特点,并分析原因。
(2)分析马达加斯加岛生物多样性丰富且独特物种较多的自然原因。
(3)从地理环境整体性的角度,分析流经刚果盆地的河流水文特征。
8 . 土壤有机质是衡量土壤肥力的主要指标。下图示意我国伊犁河谷不同植被带0~10厘米土层深度土壤有机碳含量变化及冬季雪岭云杉景观图。读图完成下面小题。
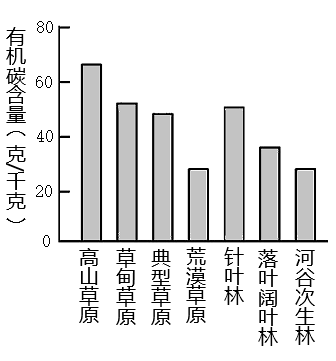

1.影响伊犁河谷山地土壤有机碳含量差异的主导因素是( )
| A.土层厚度 | B.光照 | C.海拔 | D.人类活动 |
| A.四季常绿 | B.树冠浑圆 | C.纸质叶片 | D.结构复杂 |
| A.淋溶作用强——常年径流大 | B.枯枝落叶厚——低温分解慢 |
| C.土层分层多——土层原度大 | D.土壤酸性强——矿物分解快 |
高山林线作为郁闭森林和高山植被之间的生态过渡带,是极端环境条件下树木生存的界限。林线交错带生物生存环境梯度变化剧烈。色季拉山位于雅鲁藏布江大拐弯西北侧,地处藏东南湿润气候区与半湿润气候区的分界面上,受印度洋季风的影响,其林线处森林植物群落类型呈现多样性特征。
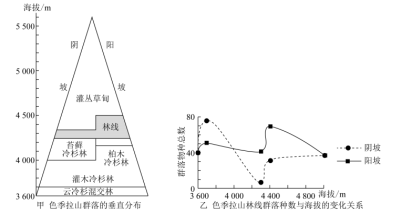
(1)简述色季拉山林线高度阳坡高于阴坡的原因。
(2)说明色季拉山阴坡林线处物种丰富度的特点及形成原因。
藤本植物是一种自身不能构成群落,必须依附它物而生的植物。植物体经常或林冠或林下或匍匐或攀援,长期生活于这种状态下的藤本植物必然形成一些特殊的行为和适应特征。一般来说,藤本植物指示着生存环境具有湿热特性。岳麓山风景区位于中国中部湖南省长沙市岳麓区(图左),位于东亚典型的季风气候区域,植被以典型的亚热带常绿阔叶林和亚热带暖性针叶林为主。当地某中学地理研学小组对岳麓山风景区的藤本植物进行考察,对山地不同方位的藤本植物数量进行统计并绘制成图(图右)。
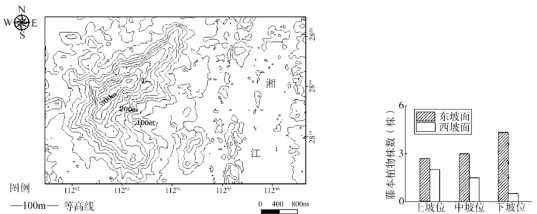
(2)描述西坡面藤本植物在不同坡位的空间分布特点,并分析其成因。
(3)与下坡位相比,东坡面上坡位藤本植物株数较少,但盖度(地上部分垂直投影的面积占地面的比率)较大。分析造成这种差异的原因。


