水稻(2n=24)是我国主要粮食作物,杂交水稻具有产量上的优势。我国学者发现了水稻雄性不育株(雄蕊发育异常,简称不育株),在杂交育种中起到重要作用。
(1)育种过程中,相比两性水稻,雄性不育株作母本的优势是___ 。
(2)育性正常与雄性不育水稻进行杂交,F1中育性正常与雄性不育各占一半,F1育性正常的自交后代全部育性正常。依据结果可知:___ 为隐性性状。有人推测雄性不育株细胞质中不存在独立控制不育性状的基因,请写出此推测的理由___ 。
(3)水稻的育性是由细胞质基因和细胞核基因共同控制的(细胞核基因:R-可育,r-不育;细胞质基因:N-可育,S-不育),只有核质基因均为不育时才表现为雄性不育,记为S(rr)。品系1(雄性不育)与品系2杂交所产子代仍表现为雄性不育,由此可大量繁殖品系1,推测品系2与育性相关的基因型为___ 。品系1与品系3(育性正常)杂交,收获所结种子S(杂合子)可供农民生产种植,推测品系3与育性相关的基因型为___ 。
(4)研究发现,光温敏不育基因tms5的突变导致水稻无法表达一种RNA水解酶,该酶能水解Ub40基因转录产生的mRNA,后者在高温下表达增强,Ub40蛋白积累导致花粉不育。科研人员利用60Co照射籼稻品种,选育了一种光温敏雄性不育品种。请设计一个实验探究新发现的光温敏不育株是tms5基因的突变还是一个新基因的突变。
实验思路:___ 。
预期结果和结论:___ 。
(1)育种过程中,相比两性水稻,雄性不育株作母本的优势是
(2)育性正常与雄性不育水稻进行杂交,F1中育性正常与雄性不育各占一半,F1育性正常的自交后代全部育性正常。依据结果可知:
(3)水稻的育性是由细胞质基因和细胞核基因共同控制的(细胞核基因:R-可育,r-不育;细胞质基因:N-可育,S-不育),只有核质基因均为不育时才表现为雄性不育,记为S(rr)。品系1(雄性不育)与品系2杂交所产子代仍表现为雄性不育,由此可大量繁殖品系1,推测品系2与育性相关的基因型为
(4)研究发现,光温敏不育基因tms5的突变导致水稻无法表达一种RNA水解酶,该酶能水解Ub40基因转录产生的mRNA,后者在高温下表达增强,Ub40蛋白积累导致花粉不育。科研人员利用60Co照射籼稻品种,选育了一种光温敏雄性不育品种。请设计一个实验探究新发现的光温敏不育株是tms5基因的突变还是一个新基因的突变。
实验思路:
预期结果和结论:
更新时间:2024-03-20 22:30:39
|
相似题推荐
非选择题-解答题
|
较难
(0.4)
【推荐1】迟发性脊椎骨骺发育不良(简称SEDL)是一类软骨发育不良遗传病。因其发病较晚(一般10岁后才出现典型症状)而很难对症状前患者进行诊断。研究者对某SEDL家系进行了调查,结果如图1(省略的家系成员均与此病无关),图中除第V代个体外其余均已成年。
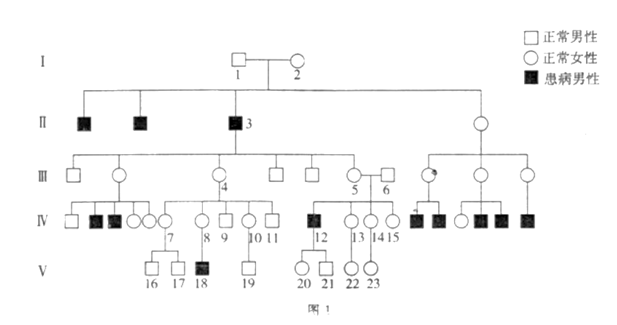
(1)据图1分析,SEDL是由___________ 性基因控制的遗传病,其致病基因最可能位于___________ 染色体上。图中第V代个体中,可以确定为杂合子的是_____________________ 。
(2)为了实现对SEDL的早期诊断和遗传预测,研究者利用遗传标记DXS16对该家系成员进行分析。已知DXS16是一类含有CA双核苷酸重复序列的标记,根据长度不同表示为D1、D2、D3……。该家系部分成员DXS16标记如图2所示。

①由图推测,遗传标记_____________ (填“D2”、“D8”或“D10”)可作为SEDL致病基因的标记。
②14、15、22号个体中,需要在孕期对胎儿进行基因诊断的是_____________ 。
③结合图1、图2,推测家系中未能提供检测样本的13号个体的DXS16标记是_____________ 。
(3)为了阐明SEDL发病的分子机制,研究人员对SEDL的致病基因和相应正常基因的结构及表达过程进行了研究。

①根据图3信息,mRNA前体加工过程中,____________ 序列均被完全剪除,一般认为它们与Sedlin蛋白的氨基酸序列不存在对应关系。
②提取患者、携带者和正常人的mRNA,经逆转录获得cDNA后PCR扩增其正常基因和致病基因,结果如下表所示。
结合图3和表中数据可知,与正常基因相比,致病基因的成熟mRNA________________ 。由于mRNA的起始密码子位于E3序列内,可推测SEDL患者发病的原因是_________________ 。
③基因测序结果表明:与正常基因相比,致病基因仅在Ⅰ2区域发生了A/T→C/G碱基对的替换,这种变异方式______________________ (填“属于”或“不属于”)基因突变。
④综合上述研究结果推测,致病基因Ⅰ2区域的碱基变化导致SEDL的原因____________ 。
(1)据图1分析,SEDL是由
(2)为了实现对SEDL的早期诊断和遗传预测,研究者利用遗传标记DXS16对该家系成员进行分析。已知DXS16是一类含有CA双核苷酸重复序列的标记,根据长度不同表示为D1、D2、D3……。该家系部分成员DXS16标记如图2所示。
①由图推测,遗传标记
②14、15、22号个体中,需要在孕期对胎儿进行基因诊断的是
③结合图1、图2,推测家系中未能提供检测样本的13号个体的DXS16标记是
(3)为了阐明SEDL发病的分子机制,研究人员对SEDL的致病基因和相应正常基因的结构及表达过程进行了研究。
①根据图3信息,mRNA前体加工过程中,
②提取患者、携带者和正常人的mRNA,经逆转录获得cDNA后PCR扩增其正常基因和致病基因,结果如下表所示。
| mRNA来源 | 患者 | 携带者 | 正常人 |
| 扩增产物长度(bp) | 567、425 | 679、567、537、425 | 679、537 |
结合图3和表中数据可知,与正常基因相比,致病基因的成熟mRNA
③基因测序结果表明:与正常基因相比,致病基因仅在Ⅰ2区域发生了A/T→C/G碱基对的替换,这种变异方式
④综合上述研究结果推测,致病基因Ⅰ2区域的碱基变化导致SEDL的原因
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
【推荐2】果蝇的眼色有多种且受多对基因控制,有色眼性状与细胞内色素的种类及积累量有关。W基因表达产物为生物膜上的色素前体物质转运载体,V基因、P基因表达产物为V酶、P酶,相关色素的合成过程如下图所示。回答下列问题:
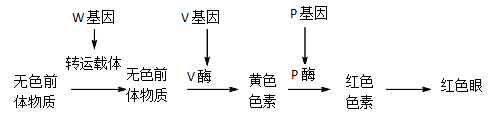
(1)据图分析,若发生单基因隐性突变会出现白眼果蝇,则发生突变的基因可能是__________ 。基因突变是指DNA分子中发生碱基对的__________ ,而引起的____________ 的改变。若该突变是由于基因中插入了一个碱基对造成的,该基因翻译时,可能由于____________ 或____________ ,产生异常的蛋白质。
(2)已知基因V/v和基因P/p位于常染色体上,基因W/w位于X染色体上。
①就上述多对等位基因而言,纯合白眼雄果蝇的基因型有____________ 种,纯合红眼雌果蝇的基因型为____________ 。
②若一对红眼果蝇杂交,F1个体表现为红眼∶黄眼∶白眼=27∶9∶28,则亲代雌果蝇的基因型是____________ ,F1中黄眼雄果蝇的基因为____________ 。
(1)据图分析,若发生单基因隐性突变会出现白眼果蝇,则发生突变的基因可能是
(2)已知基因V/v和基因P/p位于常染色体上,基因W/w位于X染色体上。
①就上述多对等位基因而言,纯合白眼雄果蝇的基因型有
②若一对红眼果蝇杂交,F1个体表现为红眼∶黄眼∶白眼=27∶9∶28,则亲代雌果蝇的基因型是
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
名校
解题方法
【推荐3】番茄花的雄蕊花药合生,包裹住雌蕊,如图1;雌蕊的柱头有3种类型:柱头外露(柱头高出雄蕊花药)、柱头齐平(柱头与雄蕊花药等高)、柱头内陷(柱头矮于雄蕊花药),如图2,野生番茄一般为柱头外露型,在种植驯化为栽培番茄时,逐步往柱头内陷型演化。
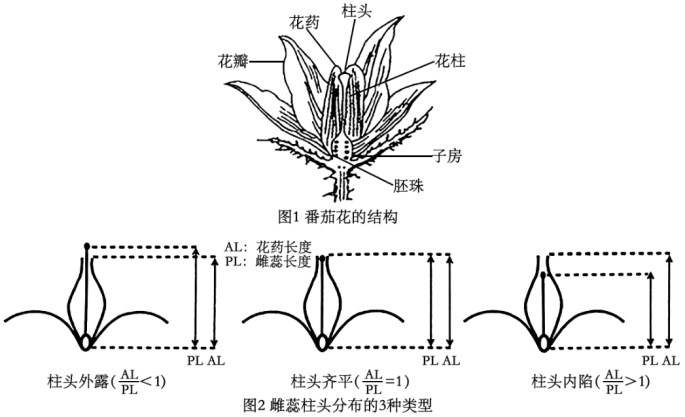
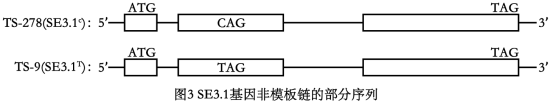
(1)为研究番茄柱头发育的遗传基础,研究人员利用柱头齐平的番茄品系(TS-278)、柱头内陷的番茄品系(TS-9)运用全基因组关联分析确认一个与柱头发育相关的基因(SE3.1),比较TS-278与TS-9的SE3.1基因非模板链的碱基序列差异如图3所示。(起始密码:AUG,终止密码:UAA、UGA、UAG)
①据图3推测,柱头内陷类型产生的原因可能是基因SE3.1发生碱基C替换成T,使____________ ,导致蛋白产物功能丧失,影响柱头发育,使栽培番茄柱头内陷于花药内部。
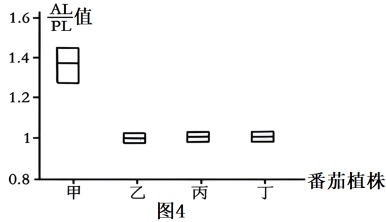
②为验证SE3.1基因对番茄柱头发育的控制功能,科研人员将SE3.1c基因导入TS-9的组织细胞中,培育出相应的转基因番茄植株,分别检测非转基因植株和转基因番茄植株AL/PL值,结果如图4,则属于转基因番茄植株的有____________ 。
(2)已有研究表明,Style2.1基因也参与影响番茄柱头发育。为了探究SE3.1基因与Style2.1基因在番茄柱头发育过程中作用特点,研究人员以柱头外露的番茄品系TS-19(具有SE3.1c基因与功能正常的Style2.1基因>为实验材料,利用基因敲除技术分别创建SE3.1基因功能缺陷的单突变株、Style2.1基因功能缺陷的单突变株、SE3.1基因和Style2.1基因功能皆缺陷的双突变株,分别检测TS-19植株及各突变株的AL/PL值,结果如图5。
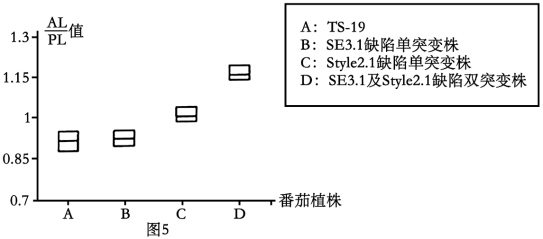
综上实验结果,请完善野生番茄柱头外露演化为栽培番茄柱头内陷的过程模型如下:

①____________________ ;
②_____________________ 。
基于上述模型推测,野生番茄被驯化为栽培番茄,其受粉方式由异化受粉转化为自花受粉,从而有助于结出更多、更大的番茄,提升产量。番茄产量提升的可能原因是番茄授粉方式转变后提高了_______________ 。
(3)为探究SE3.1基因与Style2.1基因是否独立遗传,请以TS-9、TS-19、TS-278为材料,设计遗传实验方案,并预期支持其独立遗传的结果_____________________________ 。
(1)为研究番茄柱头发育的遗传基础,研究人员利用柱头齐平的番茄品系(TS-278)、柱头内陷的番茄品系(TS-9)运用全基因组关联分析确认一个与柱头发育相关的基因(SE3.1),比较TS-278与TS-9的SE3.1基因非模板链的碱基序列差异如图3所示。(起始密码:AUG,终止密码:UAA、UGA、UAG)
①据图3推测,柱头内陷类型产生的原因可能是基因SE3.1发生碱基C替换成T,使
②为验证SE3.1基因对番茄柱头发育的控制功能,科研人员将SE3.1c基因导入TS-9的组织细胞中,培育出相应的转基因番茄植株,分别检测非转基因植株和转基因番茄植株AL/PL值,结果如图4,则属于转基因番茄植株的有
(2)已有研究表明,Style2.1基因也参与影响番茄柱头发育。为了探究SE3.1基因与Style2.1基因在番茄柱头发育过程中作用特点,研究人员以柱头外露的番茄品系TS-19(具有SE3.1c基因与功能正常的Style2.1基因>为实验材料,利用基因敲除技术分别创建SE3.1基因功能缺陷的单突变株、Style2.1基因功能缺陷的单突变株、SE3.1基因和Style2.1基因功能皆缺陷的双突变株,分别检测TS-19植株及各突变株的AL/PL值,结果如图5。
综上实验结果,请完善野生番茄柱头外露演化为栽培番茄柱头内陷的过程模型如下:
①
②
基于上述模型推测,野生番茄被驯化为栽培番茄,其受粉方式由异化受粉转化为自花受粉,从而有助于结出更多、更大的番茄,提升产量。番茄产量提升的可能原因是番茄授粉方式转变后提高了
(3)为探究SE3.1基因与Style2.1基因是否独立遗传,请以TS-9、TS-19、TS-278为材料,设计遗传实验方案,并预期支持其独立遗传的结果
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
名校
【推荐1】通过各种方法改善农作物的遗传性状,提高粮食产量一直是科学家不断努力追求的目标。如图表示一些育种途径,请回答相关问题。
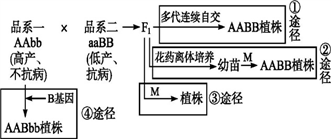
(1)①途径称为杂交育种,所利用的原理是________ ,这种育种方法除了可以得到符合要求的纯合子,还可利用植物的________ 提高产量。
(2)最常用的试剂M是________ ,该试剂能让细胞内染色体数目加倍的机理是__________________ 。相对于①途径,②途径的优势在于______________ 。
(3)可定向改变生物遗传性状的是___ 途径。经③途径所得到的植株基因型为______ 。
(4)某植物种群中基因型为AA的个体占20%,基因型为aa的个体占50%。若人为舍弃隐性性状类型仅保留显性性状类型,令其自交,则自交子一代所有个体中基因型为AA的个体占____ ,此时种群中A的基因频率为____ ,经这种人工选择作用,该种群是否发生了进化?___ 。
(1)①途径称为杂交育种,所利用的原理是
(2)最常用的试剂M是
(3)可定向改变生物遗传性状的是
(4)某植物种群中基因型为AA的个体占20%,基因型为aa的个体占50%。若人为舍弃隐性性状类型仅保留显性性状类型,令其自交,则自交子一代所有个体中基因型为AA的个体占
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
【推荐2】水稻的穗大(A)对穗小(a)为显性,晚熟(B)对早熟(b)显性,且控制这两对性状的基因独立遗传。请回答与水稻品种选育相关的问题:
(1)现有基因型为Aa的大穗水稻,A和a不影响个体生活力,且不考虑基因突变。若进行连续多代自交,并逐代淘汰隐性个体,则F2中能稳定遗传的大穗水稻占__________ 。
(2)现有品种中,大穗稻晚熟,早熟水稻穗小。为获得大穗早熟水稻新品种,某研究人员的杂交育种方案设计如下:
将杂交子一代大穗晚熟(甲)自交,得到子二代:大穗晚熟(乙)、大穗早熟(丙)、小穗晚熟(丁)、小穗早熟(戊)。单株收获F2中丙植株上的种子并单独种植,最早在子_______ 代中,能分离得到大穗早熟纯合子。若取丙植株单独种植并进行相互交配,则后代中大穗早熟的纯合子所占比例为_________ 。若想在最短时间内选育出符合要求的水稻品种,可选取上述植株____________ 的花粉进行育种。
(3)杂交水稻(杂种子一代)因具有杂种优势(高产、优质、抗性强),在农业生产中常作为种子直接利用。但种子只能用一年,需年年制种,这是因为________________________ 。
(1)现有基因型为Aa的大穗水稻,A和a不影响个体生活力,且不考虑基因突变。若进行连续多代自交,并逐代淘汰隐性个体,则F2中能稳定遗传的大穗水稻占
(2)现有品种中,大穗稻晚熟,早熟水稻穗小。为获得大穗早熟水稻新品种,某研究人员的杂交育种方案设计如下:
将杂交子一代大穗晚熟(甲)自交,得到子二代:大穗晚熟(乙)、大穗早熟(丙)、小穗晚熟(丁)、小穗早熟(戊)。单株收获F2中丙植株上的种子并单独种植,最早在子
(3)杂交水稻(杂种子一代)因具有杂种优势(高产、优质、抗性强),在农业生产中常作为种子直接利用。但种子只能用一年,需年年制种,这是因为
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
名校
【推荐3】甜瓜是世界主要食用水果之一,甜瓜是一年生植物,没有性染色体,但花朵有3种类型:雄花(♂)、雌花(♀)和全花(即两性花,一朵花中有雌蕊和雄蕊,用⚥表示)。
(1)花朵类型的任意组合,使甜瓜有______ 种性别类型。其中常见的品系是雄全同株(♂+⚥)、两性花株(⚥)。目前商业栽培的甜瓜品种为两品系杂交产生的F1具有明显的杂种优势,但在制备时需要 ______ ,工作量大,筛选全(♀)品系具有重要意义。
(2)研究者在自然界中发现了雌雄异花植株(♀+⚥),用该植株和某两性花品系(⚥)杂交,F1全部是雌雄异花植株(♀+♂),F1自交得到的F2性状分离比如下表所示。
①上述实验说明甜瓜性别是由______ 染色体上的 ______ 基因决定的。
②外施乙烯可以将雌雄异花同株(♀+♂)的雄花变成单性雌花,说明乙烯对雌蕊发育起______ 作用。研究发现基因控制乙烯合成ACC的合成,在ACC催化产生的中间产物的基础上,aa基因诱导合成赤霉素促进雄蕊的发育,基因型为aagg的植株最终发育成两性花株(⚥)。据此分析雌全同株(♀+⚥)的基因型A_gg,雄全同株(♀+⚥)的基因型为 ______ 。
(3)科研人员在雌全同株(♀+⚥)AAgg培育过程发现了全雌(♀)突变体,二者杂交后再自交,发现F2雌全同株(♀+⚥):全雌(♀)突变体分离比为3:1,则全雌对雌全同株为______ 性状。全雌(♀)突变体与常见的两种性别类型的甜瓜杂交,都可以在 ______ 植株上收获种子用于商业种植,免去了之前杂交育种的繁琐。
(4)为了源源不断获得全雌植株(♀)进行杂交育种,可将(3)F2中的雌全同株(♀+⚥)自交,在F3幼苗期扩增雌花决定基因(用GY/gy表示),然后经过AluⅠ酶切、电泳鉴定基因型,结果如图所示。

由结果可知,F3中______ 号为全雌植株(♀),将其移栽保留,另外应淘汰 ______ 号植株,原因是 ______ 。
(5)研究者发现在花芽时期,用一定浓度的赤霉素(GA3)或者硝酸银(AgNO3)可以诱导全雌植株产生一定量的两性花,从而实现自交保存。实验结果如下表所示。下表中第Ⅰ组处理试剂和浓度分别为______ 。
(6)请评价(4)和(5)中所述育种方法的优缺点______ 。
(1)花朵类型的任意组合,使甜瓜有
(2)研究者在自然界中发现了雌雄异花植株(♀+⚥),用该植株和某两性花品系(⚥)杂交,F1全部是雌雄异花植株(♀+♂),F1自交得到的F2性状分离比如下表所示。
♀+♂ | ♀+⚥ | ♂+⚥ | ⚥ |
9 | 3 | 3 | 1 |
①上述实验说明甜瓜性别是由
②外施乙烯可以将雌雄异花同株(♀+♂)的雄花变成单性雌花,说明乙烯对雌蕊发育起
(3)科研人员在雌全同株(♀+⚥)AAgg培育过程发现了全雌(♀)突变体,二者杂交后再自交,发现F2雌全同株(♀+⚥):全雌(♀)突变体分离比为3:1,则全雌对雌全同株为
(4)为了源源不断获得全雌植株(♀)进行杂交育种,可将(3)F2中的雌全同株(♀+⚥)自交,在F3幼苗期扩增雌花决定基因(用GY/gy表示),然后经过AluⅠ酶切、电泳鉴定基因型,结果如图所示。
由结果可知,F3中
(5)研究者发现在花芽时期,用一定浓度的赤霉素(GA3)或者硝酸银(AgNO3)可以诱导全雌植株产生一定量的两性花,从而实现自交保存。实验结果如下表所示。下表中第Ⅰ组处理试剂和浓度分别为
编号 | 处理苗数 | 处理试剂 | 处理浓度(mg/L) | 两性花诱导率(%) |
1 | 10 | _____ | _____ | 0 |
2 | 20 | AgNO3 | 100 | 5.56 |
3 | 20 | AgNO3 | 300 | 11.11 |
4 | 20 | AgNO3 | 500 | 27.78 |
5 | 20 | GA3 | 300 | 27.78 |
6 | 20 | GA3 | 500 | 22.23 |
7 | 20 | GA3 | 700 | 5.56 |
(6)请评价(4)和(5)中所述育种方法的优缺点
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
【推荐1】有一类玉米能够正常进行自交,它们也可以给其它类型玉米授粉结实,而其它类型玉米给它们授粉则不结实,这种现象称之为玉米单向杂交不亲和性(简称不亲系)。某不亲系受基因A控制,其遗传机制为:含有A的卵细胞与a花粉受精产生后代死亡,其余配子间结合方式均能正常结实。玉米甜与非甜性状受一对等位基因Bb控制。研究人员选择纯种不亲系甜玉米与普通的非甜玉米杂交,F1均为不亲系非甜玉米,F1与纯种普通甜玉米杂交,F2中不亲系甜玉米:普通非甜玉米=1:1。不考虑突变和染色体互换。回答下列问题:
(1)玉米甜与非甜相对性状中,隐性性状为___________ 。F1与纯种普通甜玉米杂交时,F1应作为____________ (填“父本”或“母本”),F2性状之比为1:1的原因是____________ 。若F2自交,子代的表型及比例为____________ 。
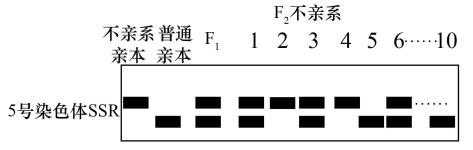
(2)SSR是DNA中的简单重复序列,非同源染色体上的SSR、不同品种的同源染色体上的SSR不同。利用SSR技术可以进行基因在染色体上的定位。研究人员为了确定A基因是否位于5号染色体上,将F1自交后提取F2中不亲系的10株玉米的叶肉细胞DNA,利用SSR进行PCR扩增,对扩增结果进行电泳,如图所示。该结果表明A基因____________ (填“在”或“不在”)5号染色体上,理由是____________ 。
(3)已知控制玉米胚乳的紫色(R)与无色(r)的基因位于9号染色体上,现有基因型为AARR与aarr玉米可供选择,请设计杂交实验验证两对基因的遗传符合自由组合定律_____ (写出实验方案及预期结果)。
(1)玉米甜与非甜相对性状中,隐性性状为
(2)SSR是DNA中的简单重复序列,非同源染色体上的SSR、不同品种的同源染色体上的SSR不同。利用SSR技术可以进行基因在染色体上的定位。研究人员为了确定A基因是否位于5号染色体上,将F1自交后提取F2中不亲系的10株玉米的叶肉细胞DNA,利用SSR进行PCR扩增,对扩增结果进行电泳,如图所示。该结果表明A基因
(3)已知控制玉米胚乳的紫色(R)与无色(r)的基因位于9号染色体上,现有基因型为AARR与aarr玉米可供选择,请设计杂交实验验证两对基因的遗传符合自由组合定律
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
名校
【推荐2】某雌雄异株植物的红花和白花为一对相对性状,受一对等位基因(位于常染色体上或仅位于X染色体上)控制。在该植物纯合红花种群中发现了一株白花雄株,研究发现该白花雄株的出现是某一亲本减数分裂时发生基因突变所导致,回答下列问题:
(1)基因突变是指______________________________________________________________________ 。
(2)若控制该相对性状的基因位于常染色体上,则该突变属于_______________ (填“显性”“隐性”或“显性或隐性”)突变,且突变基因来自______________ (填“父本”“母本”或“父本或母本”)。
(3)若控制该相对性状的基因仅位于X染色体上,则该突变属于_______________ (填“显性”“隐性”或“显性或隐性”)突变,且突变基因来自______________ (填“父本”“母本”或“父本或母本”)。
(4)请设计一次杂交实验探究控制该对相对性状的基因的位置(要求:写出实验思路,并预期结果和结论)。____________________________________________
(1)基因突变是指
(2)若控制该相对性状的基因位于常染色体上,则该突变属于
(3)若控制该相对性状的基因仅位于X染色体上,则该突变属于
(4)请设计一次杂交实验探究控制该对相对性状的基因的位置(要求:写出实验思路,并预期结果和结论)。
您最近一年使用:0次
非选择题-解答题
|
较难
(0.4)
【推荐3】科研人员用化学诱变剂处理野生型水稻,筛选得到斑点叶突变体S1和S2.为研究突变性状的遗传规律,进行了两组杂交实验,过程及结果如下表所示。
(1)两组杂交F1均为野生型,F1自交得到的F2群体中野生型与突变型的比例接近_____ ,判断S1和S2的突变性状均由一对_____ ( 显/隐)性基因控制。
(2)为研究S1和S2两种突变体的突变基因是否为同一基因突变导致,若用杂交组一的F1与杂交组二的F1作为亲本杂交,观察并统计其后代的表型。
①若子代表型为_____ ,则两种突变体的突变基因为同一基因。
②若子代表型为_____ ,则两种突变体的突变基因位于同一条染色体的不同位置上。
(3)若将第(2)问中两组F1杂交获得的所有稻粒进行单株种植并自交,单独统计每株所产生后代中的野生型和斑点叶两种表型比例。若有部分植株的后代野生型和斑点叶的比例都接近_____ ,则证明两突变体的突变基因分别位于两对同源染色体上独立遗传。
| 序号 | 杂交组合 | F1群体 | F1自交得到的F2群体(株) | ||
| 野生型 | 斑点叶 | 总数 | |||
| 杂交组一 | S1×野生型 | 野生型 | 171 | 56 | 227 |
| 杂交组二 | S2×野生型 | 野生型 | 198 | 68 | 266 |
(2)为研究S1和S2两种突变体的突变基因是否为同一基因突变导致,若用杂交组一的F1与杂交组二的F1作为亲本杂交,观察并统计其后代的表型。
①若子代表型为
②若子代表型为
(3)若将第(2)问中两组F1杂交获得的所有稻粒进行单株种植并自交,单独统计每株所产生后代中的野生型和斑点叶两种表型比例。若有部分植株的后代野生型和斑点叶的比例都接近
您最近一年使用:0次


