某哺乳动物(性别决定方式为XY型)的毛色由3对位于常染色体上、独立遗传的等位基因决定,其中A基因编码的酶1可使黄色素转化为褐色素,B基因编码的酶2可使该褐色素转化为黑色素,D基因的表达产物能完全抑制A基因的表达,相应的隐性等位基因a、b、d的表达产物没有上述功能。回答下列问题:
(1)毛色为黄色的个体的基因型有______ 种,其中纯合子的基因型有______ 种。
(2)若基因型为AaBbdd的多只雌雄动物杂交多次,产生的子代毛色及其性状分离比是______ 。若用两个纯合黄色品种的动物作为亲本进行杂交,F1均为黄色,F2中毛色表型出现了黄色∶褐色∶黑色=52∶3∶9的数量比,则杂交亲本的组合是______ 。
(3)该动物的长毛和短毛是一对相对性状,由基因E/e控制,有角和无角是一对相对性状,由基因F/f控制。研究人员让一对长毛有角的雌雄动物进行交配得到F1,其表型及比例如下表。不考虑X和Y染色体的同源区段,也不考虑互换和突变。
①基因F/f位于______ 染色体上,F1雌性个体出现表中所示比例的原因是______ 。
②多只无角雌性个体与多只有角雄性个体杂交,子代出现了一只无角雌性个体。研究人员推测可能是基因突变或染色体结构缺失造成上述现象。请你设计实验对上述推测进行验证。(注:各型配子活力相同;一对同源染色体都缺失相同片段时胚胎才致死;雄性个体缺失有角基因时存活且表现为无角。)
实验思路:________________ 。
预期实验结果:__________________________ 。
(1)毛色为黄色的个体的基因型有
(2)若基因型为AaBbdd的多只雌雄动物杂交多次,产生的子代毛色及其性状分离比是
(3)该动物的长毛和短毛是一对相对性状,由基因E/e控制,有角和无角是一对相对性状,由基因F/f控制。研究人员让一对长毛有角的雌雄动物进行交配得到F1,其表型及比例如下表。不考虑X和Y染色体的同源区段,也不考虑互换和突变。
♀ | ♂ |
短毛有角∶长毛有角=2∶5 | 短毛无角∶长毛无角∶短毛有角∶长毛有角=1∶3∶1∶3 |
②多只无角雌性个体与多只有角雄性个体杂交,子代出现了一只无角雌性个体。研究人员推测可能是基因突变或染色体结构缺失造成上述现象。请你设计实验对上述推测进行验证。(注:各型配子活力相同;一对同源染色体都缺失相同片段时胚胎才致死;雄性个体缺失有角基因时存活且表现为无角。)
实验思路:
预期实验结果:
更新时间:2024-05-25 10:01:53
|
相似题推荐
非选择题-解答题
|
困难
(0.15)
【推荐1】果蝇的翅形有有翅(长翅、短翅) 和无翅,控制翅型的两对基因A、a和B、b分别位于常染色体和X染色体上。现有甲、乙两组果蝇进行了杂交实验,其中乙组子一代雌雄果蝇随机交配得到子二代,结果如下表:
回答下列问题:
(1)果蝇的翅形性状中,________ (填有翅或无翅)是显性性状。
(2)果蝇的翅型基因中,控制________ (填有无翅或长短翅)的基因位于X染色体上,判断的依据是_______________________________ 。
(3)甲组亲本的基因型为_____________ ,若让甲组F1的长翅果蝇随机交配,则F2雄果蝇中,长翅∶短翅∶无翅=________________ 。
(4)选择甲组F1无翅雄果蝇与乙组F2的长翅雌果蝇杂交,其子代中长翅雌果蝇所占的比例为___________ 。
(5)欲通过一代杂交实验鉴定某短翅雌果蝇的基因型,可选择表现型为________ 的雄果蝇与其杂交,若后代雄果蝇的翅形均表现为__________ ,则该雌果蝇为纯合子。
杂交组别 | 亲本(P) | 子一代(F1) ♀:♂=1:1 | 子二(F2)♀:♂=1:1 | |||
♀ | ♂ | ♀ | ♂ | ♀ | ♂ | |
甲组 | 长翅 | 无翅 | 长翅:无翅 =1∶1 | 长翅:短翅:无翅 =1∶1∶2 | - | - |
乙组 | 短翅 | 无翅 | 全为长翅 | 全为短翅 | 长翅:短翅:无翅 =3∶3∶2 | 长翅:短翅:无翅=3∶3∶2 |
(1)果蝇的翅形性状中,
(2)果蝇的翅型基因中,控制
(3)甲组亲本的基因型为
(4)选择甲组F1无翅雄果蝇与乙组F2的长翅雌果蝇杂交,其子代中长翅雌果蝇所占的比例为
(5)欲通过一代杂交实验鉴定某短翅雌果蝇的基因型,可选择表现型为
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
名校
【推荐2】某二倍体昆虫的翅型和眼型分别受两对等位基因Aa、Bb控制(两对基因均不在Y染色体上,且其中一对性状存在某种基因型致死现象)。为研究其遗传机制,让一只长翅正常眼雌性昆虫与一只短翅正常眼雄性个体交配,F1中的雌性个体全为正常眼,雄性个体中有正常眼和细眼,让F1中的雌雄个体相互交配,F2的表现型及比例如下表:
(1)Aa、Bb两对等位基因的遗传符合______ 定律。
(2)两对等位基因中,位于X染色体上的是_____ ,翅型中显性性状为______ 。
(3)亲代雌雄个体的基因型为_______ 。致死基因型为_______ 。
(4)让F2中所有短翅个体随机交配,理论上F3的短翅雌性昆虫中纯合个体所占比例为_______ 。
(5)为进一步验证致死基因型,可选用F1中的长翅正常眼雄性个体与F2中长翅正常眼雌性个体进行杂交,用遗传图解预测实验结果______________ 。
| 短翅正常眼 | 短翅细眼 | 长翅正常眼 | 长翅细眼 | |
| 雌性个体 | 1/8 | 1/16 | 9/56 | 9/112 |
| 雄性个体 | 1/8 | 1/8 | 9/56 | 9/56 |
(1)Aa、Bb两对等位基因的遗传符合
(2)两对等位基因中,位于X染色体上的是
(3)亲代雌雄个体的基因型为
(4)让F2中所有短翅个体随机交配,理论上F3的短翅雌性昆虫中纯合个体所占比例为
(5)为进一步验证致死基因型,可选用F1中的长翅正常眼雄性个体与F2中长翅正常眼雌性个体进行杂交,用遗传图解预测实验结果
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
【推荐3】如下图所示,某二倍体植物有红花和白花两种类型,受三对等位基因控制(三对基因独立遗传,用A、a,B、b和D、d表示)。回答下列问题:
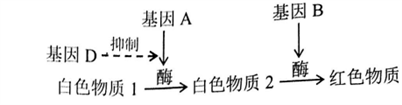
(1)据图可知,基因通过____________ ,进而控制该植物的花色,红花植株的基因型有____________ 种。
(2)某红花植株自交,子—代的性状分离比为3:1,则该红花植株的基因型为____________ 。
(3)研究发现,当体细胞中其它基因数量与染色体均正常时,若b基因数多于B,B基因的表达会减弱而形成粉红花突变体,其基因与染色体的组成可能出现下图所示的三种情况:

突变体____________ 属于染色体结构变异。若不考虑基因突变和交叉互换,突变体I产生b配子的概率为____________ ;突变体II能形成B、bb两种概率相等的配子,原因是____________ ;用基因型AaBbbdd的粉花植株与基因型AAbbdd的植株杂交;若子代表现型及比例为____________ ,则该粉花植株属于突变体III类型。
(1)据图可知,基因通过
(2)某红花植株自交,子—代的性状分离比为3:1,则该红花植株的基因型为
(3)研究发现,当体细胞中其它基因数量与染色体均正常时,若b基因数多于B,B基因的表达会减弱而形成粉红花突变体,其基因与染色体的组成可能出现下图所示的三种情况:
突变体
您最近一年使用:0次
非选择题-实验题
|
困难
(0.15)
名校
【推荐1】研究人员发现了一种新的亮红眼突变型果蝇,为探究亮红眼基因突变体的形成机制,设计了一系列实验。
(1)亮红眼突变型果蝇与野生型果蝇进行_______ (填字母:a正交和反交,b自交,c测交)实验后,F1均为野生型,F2野生型与亮红眼表现型比为3:1,亮红眼果蝇雌雄个体数相当,说明亮红眼是一种位于__________ 染色体上的__________ 突变。
(2)红眼突变型果蝇还有朱红眼、朱砂眼和猩红眼等类型,朱红眼(a)、朱砂眼(b)和猩红眼(d)三个基因分别位于2号、X和3号染色体上,为探究亮红眼突变基因(用字母E或e表示)与上述三种基因的关系,以四种突变型果蝇为亲本进行杂交实验,结果如下表所示。
亮红眼与朱红眼果蝇杂交,F2性状分离比接近于9:7,可知控制亮红眼与朱红眼的基因位于两对同源染色体上,遵循自由组合定律。
①亮红眼与朱砂眼果蝇杂交,F1雄果蝇的基因型为__________ 。
②亮红眼与猩红眼果蝇杂交,F1、F2果蝇中没有出现野生型,则可以推测亮红眼基因与猩红眼基因的关系是__________ 。
(3)色氨酸是动物无法自身合成的氨基酸,称为必需氨基酸。果蝇的眼色与色素合成细胞产生的眼黄素有关,眼黄素由色氨酸经过酶促反应合成。研究发现亮红眼果蝇眼睛中眼黄素显著偏低,而色氨酸酶促反应途径没有受到影响。由此推测,亮红眼基因与色氨酸__________ 有关。
(4)相对野生型红眼果蝇而言,白眼、朱红眼均为隐性突变性状,基因均位于X染色体上。为判断三种影响眼色的突变是否为染色体同一位点的基因突变,实验过程和结果如下。
实验:P白眼♀蝇×朱红眼♂蝇 F1 红眼♀蝇:白眼♂蝇=1:1;
F1 红眼♀蝇:白眼♂蝇=1:1;
F1♀、♂相互交配 F2。
①F2出现红眼雄性的原因__________ 。
②F2雄性个体只有3%表现为红眼。由此判断,F1雌性果蝇在减数分裂形成卵细胞时,约有_____ %的初级卵细胞在这两个眼色基因位点之间发生了1次交换。
(1)亮红眼突变型果蝇与野生型果蝇进行
(2)红眼突变型果蝇还有朱红眼、朱砂眼和猩红眼等类型,朱红眼(a)、朱砂眼(b)和猩红眼(d)三个基因分别位于2号、X和3号染色体上,为探究亮红眼突变基因(用字母E或e表示)与上述三种基因的关系,以四种突变型果蝇为亲本进行杂交实验,结果如下表所示。
杂交 后代 | 亮红眼♂×朱红眼♀ | 亮红眼♂×朱砂眼♀ | 亮红眼♂×猩红眼♀ | |||
野生型 | 突变型 | 野生型 | 突变型 | 野生型 | 突变型 | |
F1 | 57♂:66♀ | 0 | 77♀ | 63♂ | 0 | 114♂:110♀ |
F2 | 116♂:118♀ | 90♂:92♀ | 75♂:79♀ | 110♂:109♀ | 0 | 227♂:272♀ |
亮红眼与朱红眼果蝇杂交,F2性状分离比接近于9:7,可知控制亮红眼与朱红眼的基因位于两对同源染色体上,遵循自由组合定律。
①亮红眼与朱砂眼果蝇杂交,F1雄果蝇的基因型为
②亮红眼与猩红眼果蝇杂交,F1、F2果蝇中没有出现野生型,则可以推测亮红眼基因与猩红眼基因的关系是
(3)色氨酸是动物无法自身合成的氨基酸,称为必需氨基酸。果蝇的眼色与色素合成细胞产生的眼黄素有关,眼黄素由色氨酸经过酶促反应合成。研究发现亮红眼果蝇眼睛中眼黄素显著偏低,而色氨酸酶促反应途径没有受到影响。由此推测,亮红眼基因与色氨酸
(4)相对野生型红眼果蝇而言,白眼、朱红眼均为隐性突变性状,基因均位于X染色体上。为判断三种影响眼色的突变是否为染色体同一位点的基因突变,实验过程和结果如下。
实验:P白眼♀蝇×朱红眼♂蝇
F1 红眼♀蝇:白眼♂蝇=1:1;F1♀、♂相互交配
F2。①F2出现红眼雄性的原因
②F2雄性个体只有3%表现为红眼。由此判断,F1雌性果蝇在减数分裂形成卵细胞时,约有
您最近一年使用:0次
非选择题-实验题
|
困难
(0.15)
【推荐2】小鼠体因体型小、易饲养、易繁殖等优点,是遗传学常用的实验材料之一。请分析回答:
(1)已知小鼠的黄毛和灰毛受一对等位基因控制。一只黄毛雌鼠与一只灰毛雄鼠杂交,F1的雌、雄中均既有黄毛,又有灰毛。请利用F1中的小鼠为材料,设计两个一代杂交实验均能独立证明控制黄毛的基因位于X染色体上,并表现为显性。(要求:请写出杂交组合和预期的实验结果,用表现型表示。)______
(2)基因型AaXBY的雄鼠(两对基因位于两对非同源染色体上),其某个精原细胞产生了一个基因组成为AAaXB的精子,请你给出合理的解释____________ 。
(1)已知小鼠的黄毛和灰毛受一对等位基因控制。一只黄毛雌鼠与一只灰毛雄鼠杂交,F1的雌、雄中均既有黄毛,又有灰毛。请利用F1中的小鼠为材料,设计两个一代杂交实验均能独立证明控制黄毛的基因位于X染色体上,并表现为显性。(要求:请写出杂交组合和预期的实验结果,用表现型表示。)
(2)基因型AaXBY的雄鼠(两对基因位于两对非同源染色体上),其某个精原细胞产生了一个基因组成为AAaXB的精子,请你给出合理的解释
您最近一年使用:0次
非选择题-实验题
|
困难
(0.15)
【推荐3】果蝇因其体积小,繁殖快等优点成为常见的遗传学实验材料。现有科研人员研究发现果蝇的黑身、灰身由一对等位基因(A、a)控制。另一对同源染色体上的等位基因(B、b)会影响黑身果蝇的体色深度。科研人员进行了以下两个实验:
实验一:黑身雌蝇甲与灰身雄蝇乙杂交,F1全为灰身,F1随机交配,F2雌雄果蝇表现型比均为灰身:黑身=3:1。
实验二:黑身雌蝇丙(基因型同甲)与灰身雄蝇丁杂交,F1全为灰身,F1随机交配,F2表现型比为:雌蝇中灰身:黑身=3:1;雄蝇中灰身:黑身:深黑身=6:1:1。
(1)A、a基因位于___________ 染色体上,若将实验一中所有果蝇混合培养,让其随机交配,子一代中有8400只灰身果蝇和1600只黑身果蝇,如果继续进行随机交配,则子二代中Aa的基因型频率为________ 。
(2)B、b基因位于________ 染色体上,实验二F2中灰身雄蝇共有________ 种基因型,其中AAXBY所占的比例是___________ 。
(3)实验中偶然出现一只深黑身雌蝇(染色体组型完整),请用遗传图解写出该雌蝇与实验二F1中灰身雄蝇的杂交过程(要求写出配子)。___________
实验一:黑身雌蝇甲与灰身雄蝇乙杂交,F1全为灰身,F1随机交配,F2雌雄果蝇表现型比均为灰身:黑身=3:1。
实验二:黑身雌蝇丙(基因型同甲)与灰身雄蝇丁杂交,F1全为灰身,F1随机交配,F2表现型比为:雌蝇中灰身:黑身=3:1;雄蝇中灰身:黑身:深黑身=6:1:1。
(1)A、a基因位于
(2)B、b基因位于
(3)实验中偶然出现一只深黑身雌蝇(染色体组型完整),请用遗传图解写出该雌蝇与实验二F1中灰身雄蝇的杂交过程(要求写出配子)。
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
【推荐1】某XY型性别决定的二倍体植物,其高茎与矮茎由等位基因B、b控制,红果和黄果由等位基因D、d控制,两对基因均不位于Y染色体上。现有一个矮茎植株种群,其中雌株均为红果,该植株种群的雌雄个体自由交配,F₁的表现型及比例如表所示(仅某一性状存在致死现象)。
(1)该植株红果基因突变为黄果基因属于_______________ 突变(填“显性”或“隐性”)。红果基因能突变为黄果基因,黄果基因也能突变成红果基因,说明基因突变具有_______________ 的特点。
B/b和D/d这两对等位基因符合孟德尔自由组合定律,理由是_____________________________ 。
(2)测序结果表明,突变基因b的mRNA编码氨基酸序列第373位碱基改变,由5`-CGGAG-3`变为5'CGCAG3',导致第_______________ 位氨基酸改变为_______________ 。(部分密码子及对应氨基酸:GAG谷氨酸;AGA精氨酸;GAC天冬氨酸;ACA苏氨酸;CAG谷氨酰胺)
(3)亲本雄性植株的基因型及比例为_______________ ,矮茎植株种群杂交出现表中高茎矮茎特殊比例的原因是_______________ 。若将F₁中的黄果雌株去除后,让剩余的植株自由交配。F₂雌株中,高茎红果雌株所占的比例为_______________ 。
(4)选取F₁中,矮茎黄果雌株与矮茎红果雄株杂交。请写出F₁杂交产生F₂的遗传图解(棋盘法)_______________
高茎红果 | 高茎黄果 | 矮茎红果 | 矮茎黄果 | |
雌株 | 6/42 | 1/42 | 12/42 | 2/42 |
雄株 | 5/42 | 2/42 | 10/42 | 4/42 |
B/b和D/d这两对等位基因符合孟德尔自由组合定律,理由是
(2)测序结果表明,突变基因b的mRNA编码氨基酸序列第373位碱基改变,由5`-CGGAG-3`变为5'CGCAG3',导致第
(3)亲本雄性植株的基因型及比例为
(4)选取F₁中,矮茎黄果雌株与矮茎红果雄株杂交。请写出F₁杂交产生F₂的遗传图解(棋盘法)
您最近一年使用:0次
非选择题-实验题
|
困难
(0.15)
名校
【推荐2】某雄性动物的基因型为BbXdY(2N=16)。将该动物体内一个精原细胞(DNA被32P全部标记)放在不含32P的培养液中培养,经若干次分裂后,分别用与基因B、b、D、d特异性结合的探针B(黄色)、探针b(绿色)、探针D(红色)、探针d(蓝色)处理某一分裂时期的细胞,结果如图1所示,已知该细胞在分裂过程中只发生了一次变异。回答下列问题: ___________ 分裂的后期,理由是__________________ 。___________ (填“能”或“不能”)确定该细胞中含有32P的染色体数量。图1细胞中出现红色的原因是______________________________ 。
(2)若该动物一个精原细胞中基因B和b所在染色体片段发生互换,则经探针处理后,不考虑其他变异,产生的配子中只显黄色的比例为___________ ;若该精原细胞先进行一次有丝分裂,再进行一次减数分裂,则产生的子细胞中含32P的细胞所占的比例范围是___________ 。
(3)研究发现,细胞分裂过程中p53基因的表达产物能在细胞核内调控转录因子的活性,进而抑制细胞恶性增殖。为探究熊胆对皮肤癌细胞中p53基因表达的影响,研究人员分别提取经BB(熊胆粉)处理后的皮肤癌HaCaT、KUMA6细胞株的总蛋白,采用琼脂糖凝胶电泳法分离出p53,电泳结果如图2所示。
①癌细胞容易在体内分散和转移的原因是_______________ ;推测p53基因属于__________ 基因。
②琼脂糖凝胶电泳法分离p53时,p53在电场中的迁移速率主要与_____________ 有关。由电泳结果可得出的结论是____________________ 。


(2)若该动物一个精原细胞中基因B和b所在染色体片段发生互换,则经探针处理后,不考虑其他变异,产生的配子中只显黄色的比例为
(3)研究发现,细胞分裂过程中p53基因的表达产物能在细胞核内调控转录因子的活性,进而抑制细胞恶性增殖。为探究熊胆对皮肤癌细胞中p53基因表达的影响,研究人员分别提取经BB(熊胆粉)处理后的皮肤癌HaCaT、KUMA6细胞株的总蛋白,采用琼脂糖凝胶电泳法分离出p53,电泳结果如图2所示。
①癌细胞容易在体内分散和转移的原因是
②琼脂糖凝胶电泳法分离p53时,p53在电场中的迁移速率主要与
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
解题方法
【推荐3】某个品种的牵牛花野生型植株花色为红色,在种植的过程中出现4个各不相同的白花隐性突变品系,分别用M1,M2,M3,M4表示,突变基因都位于Ⅱ号染色体上。为了研究这些突变基因的相对位置关系,对4个白花品系进行了两两杂交实验,结果如下:
(1)根据杂交的结果推断,牵牛花的花色不是由同源染色体上一对等位基因所控制的,理由是______ 。
(2)若四个白花突变品系的形成都是Ⅱ号染色体上一对等位基因改变的结果,可推断花色性状至少由______ 对等位基因所控制。野生型基因用“+”表示,M1、M2、M3、M4隐性突变基因分别用X1、X2、X3、X4表示,若已知M1突变品系的突变型基因和野生型基因之间的相对位置如左图所示,则X4的位置在______ (选填下图中的数字)。

(3)科研人员进一步研究后发现M1和M3突变体是与花色决定有关的2个基因(JSC和JXY)中的一个发生单个碱基对替换的隐性突变,对M1和M3进行相关基因的测序后得到以下结果:
注:测序结果只给出基因一条链(转录模板链)的碱基序列,如【123A/A 710G/G】表示两条同源染色体上JSC基因转录模板链的第123位碱基分别为A和A,第710位碱基分别位G和G。
由图可知M1突变体发生突变的基因是______ (填“JSC基因”或“JXY基因”),推测表格中“?”处测序结果为【______ 】。若野生型植株两条同源染色体(Ⅱ号)上各有一个基因发生突变后形成了M5和M6两个植株,相关基因测序结果如表中所示,两个植株杂交,后代的中双突变基因的个体的比例为______ (不考虑其它变异)。
| P M1×M2 M1×M3 M1×M4 M2×M3 M2×M4 M3×M4 ↓ ↓ ↓ ↓ ↓ ↓ F1 突变型 野生型 野生型 野生型 野生型 突变型 |
(1)根据杂交的结果推断,牵牛花的花色不是由同源染色体上一对等位基因所控制的,理由是
(2)若四个白花突变品系的形成都是Ⅱ号染色体上一对等位基因改变的结果,可推断花色性状至少由
(3)科研人员进一步研究后发现M1和M3突变体是与花色决定有关的2个基因(JSC和JXY)中的一个发生单个碱基对替换的隐性突变,对M1和M3进行相关基因的测序后得到以下结果:
个体表现型 | JSC基因测序结果 | JXY基因测序结果 |
M1突变体 | 【123A/A 710G/G】 | 【302G/G 560A/A】 |
M3突变体 | 【123A/A 710C/C】 | ? |
野生型 | 【123A/A 710G/G】 | 【302C/C 560A/A】 |
M5(野生型) | 【123A/G 710G/G】 | 【302C/C 560A/A】 |
M6(野生型) | 【123A/A 710G/G】 | 【302G/C 560A/A】 |
由图可知M1突变体发生突变的基因是
您最近一年使用:0次
非选择题-实验题
|
困难
(0.15)
【推荐1】正常的水稻(雌雄同株)体细胞染色体数为2n=24。现有一种三体水稻,细胞中7号染色体的同源染色体有三条,即染色体数为2n+1=25。下图为该三体水稻细胞及其产生的配子类型和比例示意图(6、7为染色体标号;A为抗病基因,a为非抗病基因;①~④为四种类型配子)。已知染色体数异常的配子(如①、③)中雄配子不能参与受精作用,其他配子均能参与受精作用且个体存活。请回答:
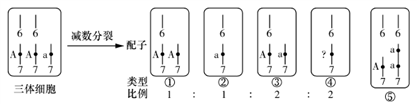
(1)图中“?”处的基因是________ ,若减数分裂过程没有发生基因突变和染色体交叉互换,则配子②和④________ (填“可能”或“不可能”)来自一个初级精母细胞。
(2)有一些抗病水稻 ,其中有部分杂合子,现通过连续自交并每代拔除感病植株来提高纯合抗病植株的比例。若该过程中,产生了一些7号染色体的三体植株,这些三体植株的存在不会影响育种目的,因为该过程中随着感病植株被拔除,
,其中有部分杂合子,现通过连续自交并每代拔除感病植株来提高纯合抗病植株的比例。若该过程中,产生了一些7号染色体的三体植株,这些三体植株的存在不会影响育种目的,因为该过程中随着感病植株被拔除,________ 不断降低;为了进一步简化操作,拟除去套袋环节,改为每代自然传粉并拔除感病植株,也可以达到提高纯合抗病植株比例的目的,但这种方法相对于连续自交来说,育种进程将________ 。(填“加快”或“减慢”或“不变”)。
(3)现有一株基因型为Aaa的抗病植株,可能是三体植株(假说1);也可能是如⑤所示由于________ 导致的(假说2)。请设计一最简捷交配实验方案(设假说2中产生的各种配子均能受精且后代均能存活),探究何种假说成立。(写出实验设计思路,预期结果和结论)
实验思路:____________________________________________________
预期结果和结论:____________________________________________________
(1)图中“?”处的基因是
(2)有一些抗病水稻
,其中有部分杂合子,现通过连续自交并每代拔除感病植株来提高纯合抗病植株的比例。若该过程中,产生了一些7号染色体的三体植株,这些三体植株的存在不会影响育种目的,因为该过程中随着感病植株被拔除,(3)现有一株基因型为Aaa的抗病植株,可能是三体植株(假说1);也可能是如⑤所示由于
实验思路:
预期结果和结论:
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
【推荐2】家蚕为ZW型性别决定的重要的模式生物,在实验中研究者偶然获得一种新的单基因纯合突变品系N(表现为第2眼纹全黑),与已报道的隐性纯合突变体M(表现为眼纹全黑,突变基因位于15号常染色体上)的表型有明显差异,为研究突变基因的定位,研究者对家蚕眼纹性状进行了遗传分析,结果如下表所示。请分析回答下列问题:
(1)分析第1组杂交实验初步判断第2眼纹全黑性状的遗传方式为____________ 。
(2)分析第2组杂交实验,控制N和M眼纹性状的基因互为____________ (填“等位基因”或“非等位基因”);据F2表型推测,它们位于____________ (填“同源染色体”或“非同源染色体”)上。F2中未出现新的突变体,其原因可能是____________ 。若将第2组实验中的F1与突变体M杂交,则子代的表型及比例为____________ 。
(3)雄蚕具有食桑量低、蚕茧率高的特点,故生产中饲养雄蚕具有更高价值。科研人员利用射线处理常染色体上控制卵色的基因(E控制黑卵,e控制白卵),得到变异家蚕,进而利用变异家蚕培育限性黑卵家蚕(如下图):

①途径X是将变异家蚕(基因型为EOZWE)与普通白卵家蚕(基因型为eeZZ)杂交得变异雌蚕,再用该变异家蚕与表型为____________ 的雄蚕杂交,最大概率地得到限性黑卵家蚕。培育限性黑卵雌蚕育种过程利用的原理是____________ 和____________ 。
②用限性黑卵家蚕与纯合的普通黑卵家蚕杂交,F1随机交配得F2,再利用专门的光电机器分选出两种不同颜色的卵。F2卵中分选出____________ (填“黑”或“白”)色卵,经孵化培养即得生产用雄蚕。已知位于W染色体上的E基因会导致常染色体上含有E的卵细胞约50%死亡,故F2中符合育种目标的卵约占总数的____________ 。
组别 | 亲本 | F1 | F2 |
第1组 | 正常眼纹♂×N♀或正常眼纹♀×N♂ | 全为正常眼纹 | 正常眼纹:第2眼纹全黑≈3:1 |
第2组 | M♂×N♀或M♀×N♂ | 全为正常眼纹 | 正常眼纹:眼纹全黑:第2眼纹全黑≈2:1:1 |
(2)分析第2组杂交实验,控制N和M眼纹性状的基因互为
(3)雄蚕具有食桑量低、蚕茧率高的特点,故生产中饲养雄蚕具有更高价值。科研人员利用射线处理常染色体上控制卵色的基因(E控制黑卵,e控制白卵),得到变异家蚕,进而利用变异家蚕培育限性黑卵家蚕(如下图):
①途径X是将变异家蚕(基因型为EOZWE)与普通白卵家蚕(基因型为eeZZ)杂交得变异雌蚕,再用该变异家蚕与表型为
②用限性黑卵家蚕与纯合的普通黑卵家蚕杂交,F1随机交配得F2,再利用专门的光电机器分选出两种不同颜色的卵。F2卵中分选出
您最近一年使用:0次
非选择题-解答题
|
困难
(0.15)
【推荐3】药用植物板蓝根(2n=14)具有抗病毒及抗菌等作用,油菜是重要的经济作物,但容易受病菌感染。某课题组将“华双3号”油菜(2n=38)与板蓝根进行体细胞融合,以期培育获得抗病菌的油菜杂交种。(图中AK5/AK6/AK8为板蓝根祖先染色体编号)

(1)研究发现,板蓝根在进化过程中发生了如图所示的变异,其中涉及的变异类型包括__________。
(2)下列如图所示的变异与板蓝根进化的关系说明,合理的是________。
根肿菌是常见的油菜病原菌。研究发现有些油菜品种具有两对独立遗传的根肿菌抗性基因。为研究抗性基因的遗传规律,以抗性植株和易感植株作为亲本进行杂交,结果如表所示。
(3)根肿菌和油菜细胞属于不同的细胞类型,其主要区别是________ 。
(4)若用A/a、B/b表示这两对基因,据表可推测,F2中易感植株的基因型共有_________ 种。抗根肿菌油菜的基因型可表示为________ 。
(5)根肿菌的种类繁多,为鉴定油菜品种感染的根肿菌类别,应________ 。(编号选填并排序)
①选取单菌落接种在液体培养基上,而后感染油菜
②从感病植物根部提取菌样制成悬液
③稀释菌液,涂布在固体培养基中
④对单个菌落进行鉴定
该课题组获得的多个体细胞杂交种中,部分品种体细胞染色体组成以及分裂过程中细胞两极染色体数目占比如表所示。
(6)在“杂交种”的育种过程中,需要________ 。(编号选填并排序)
①更换不同激素配比的培养基,诱导形成根或芽
②利用纤维素酶制备油菜和板蓝根的原生质体
③诱导原生质体形成愈伤组织
④添加促融剂促进细胞融合
(7)如表所示杂交种AS1在后期Ⅰ时,有10.6%的细胞两极染色体数目比为28:24,造成这一现象的可能原因是________ 。(编号选填)
①间期Ⅰ复制DNA出现差错
②后期Ⅰ部分同源染色体未分离
③后期Ⅱ部分姐妹染色单体未分离
④前期Ⅰ姐妹染色单体没有发生互换
(8)若要将获得的AS1、AS6和AS7等体细胞杂交种作为有性杂交的亲本,更适合选择_________ ,理由是____________ 。
(9)研究人员对“体细胞融合”策略进行改进,培育出了只含一条板蓝根染色体的“板蓝根油菜”,且这条染色体上携带有根肿病抗性基因。这一改进的优势是___________。
(1)研究发现,板蓝根在进化过程中发生了如图所示的变异,其中涉及的变异类型包括__________。
| A.易位 | B.倒位 | C.缺失 | D.重复 |
| A.Q以及X区域的保留是自然选择的结果 |
| B.V区域的基因序列改变,为进化提供丰富原材料 |
| C.板蓝根进化过程中发生了染色体结构和数目的变异 |
| D.板蓝根相比其祖先因基因数量减少,因而环境适应性降低 |
根肿菌是常见的油菜病原菌。研究发现有些油菜品种具有两对独立遗传的根肿菌抗性基因。为研究抗性基因的遗传规律,以抗性植株和易感植株作为亲本进行杂交,结果如表所示。
| 子代 | 表型数量(株) | 预期比值 | |
| 易感 | 抗根肿菌 | ||
| F1 | 91 | 1 | |
| F2 | 436 | 31 | 15:1 |
(4)若用A/a、B/b表示这两对基因,据表可推测,F2中易感植株的基因型共有
(5)根肿菌的种类繁多,为鉴定油菜品种感染的根肿菌类别,应
①选取单菌落接种在液体培养基上,而后感染油菜
②从感病植物根部提取菌样制成悬液
③稀释菌液,涂布在固体培养基中
④对单个菌落进行鉴定
该课题组获得的多个体细胞杂交种中,部分品种体细胞染色体组成以及分裂过程中细胞两极染色体数目占比如表所示。
| 杂交种 | 体细胞染色体数 | 后期I时细胞两极染色体数目比的占比(%) | ||||
| 26:26 | 28:24 | 23:29 | 27:25 | 其它 | ||
| AS1 | 52 | 63.4 | 10.6 | 7.2 | 14.8 | 0.0 |
| AS6 | 52~62 | 32.3 | 9.6 | 8.2 | 11.0 | 38.8 |
| AS7 | 52~58 | 40.3 | 9.6 | 7.2 | 12.5 | 30.5 |
①更换不同激素配比的培养基,诱导形成根或芽
②利用纤维素酶制备油菜和板蓝根的原生质体
③诱导原生质体形成愈伤组织
④添加促融剂促进细胞融合
(7)如表所示杂交种AS1在后期Ⅰ时,有10.6%的细胞两极染色体数目比为28:24,造成这一现象的可能原因是
①间期Ⅰ复制DNA出现差错
②后期Ⅰ部分同源染色体未分离
③后期Ⅱ部分姐妹染色单体未分离
④前期Ⅰ姐妹染色单体没有发生互换
(8)若要将获得的AS1、AS6和AS7等体细胞杂交种作为有性杂交的亲本,更适合选择
(9)研究人员对“体细胞融合”策略进行改进,培育出了只含一条板蓝根染色体的“板蓝根油菜”,且这条染色体上携带有根肿病抗性基因。这一改进的优势是___________。
| A.获得了双亲的大部分性状 | B.便于鉴定不同抗性基因的功能 |
| C.获得的杂交种的表型更加多样 | D.降低不同物种间基因表达的干扰程度 |
您最近一年使用:0次


