1 . 截至2023年,贵州高速公路通车里程达到8784公里。高速公路修建过程中的生物护坡绿化是公路建设工程中的一个重要组成部分,为筛选出高等级公路护坡的适宜植物,科研人员对贵州几种野生草本植物进行了引种试验,部分结果如下表。
(1)科研人员可采用______ 法调查某野地草坪百喜草种群密度,调查时关键要做到______ 。
(2)通过生物护坡绿化,既可美化环境,又可防止水土流失等,这体现了生物多样性的______ 价值。
(3)生物护坡绿化成功后,随着时间的推移,护坡区的群落被另一群落所取代,从演替的起始条件看,该过程属于______ 演替。
(4)据表分析,用于高速公路生物护坡绿化的首选草种是______ ,理由是______ 。
种名 | 青绿期(d) | 侵占性 | 再生性 | 覆盖率(%) | 综合抗逆性 | 蔓延性 |
百喜草 | 227 | 强 | 强 | 100 | 极强 | 强 |
扁穗牛鞭草 | 270 | 强 | 强 | 100 | 极强 | 强 |
宿根画眉草 | 180 | 强 | 强 | 75 | 强 | 弱 |
小颖羊茅 | 无明显枯黄 | 强 | 强 | 80 | 弱 | 弱 |
(2)通过生物护坡绿化,既可美化环境,又可防止水土流失等,这体现了生物多样性的
(3)生物护坡绿化成功后,随着时间的推移,护坡区的群落被另一群落所取代,从演替的起始条件看,该过程属于
(4)据表分析,用于高速公路生物护坡绿化的首选草种是
您最近一年使用:0次
2 . 为调查某地高原酚鼠(善于挖洞,昼伏夜出)的种群密度,科研人员开展如下工作。(1)随机选择3个0.25hm2的样方;(2)确定有效洞口:在样方内寻找新鲜的高原酚鼠土丘,挖掘其洞道,敞开洞口24h后观察,若该洞口被酚鼠重新推土堵住,则该洞口为有效洞口,统计并记录数据;(3)在样方内随机选择20个有效洞口,安放捕鼠装置,连续数日共捕获高原勖鼠7只(洞口系数=7÷20=0.35);(4)统计有关数据,计算高原酚鼠种群密度。调查数据如下表,下列相关叙述正确的是( )
某地高原勖鼠种群密度调查结果
注:洞口系数=捕获的高原鼢鼠数量/安放捕鼠装置的有效洞口数;种群密度/(只/hm2)=洞口系数×有效洞口数。
某地高原勖鼠种群密度调查结果
样地序号 | 样方面积/hm2 | 挖掘洞口/个 | 有效洞口/个 | 洞口系数 |
1 | 0.25 | 35 | 31 | 0.35 |
2 | 0.25 | 42 | 33 | 0.35 |
3 | 0.25 | 34 | 21 | 0.35 |
| A.在自然界中,高原酚鼠种群数量可呈现“J”形增长 |
| B.高原酚鼠平均有效洞口数约为113.3个/hm2,平均种群密度约为39.7只/hm2 |
| C.若某些捕鼠装置失效,则会造成估测数值比实际种群密度偏大 |
| D.高原酚鼠善于挖洞、穴居,能够改善土壤条件,增加本地原生植物产量,这体现了生物多样性的直接价值 |
您最近一年使用:0次
3 . 近段时间,广西河池市环江毛南族自治县的木论国家级自然保护区管理中心设置在保护区的红外相机,拍摄到了国家二级保护动物白鹇、猕猴等多种野生动物的珍贵影像。下列有关叙述错误的是( )
| A.还可以用逐个计数法调查部分野生动物种群数量 |
| B.白鹇、猕猴可能会与其他动物存在一定的生态位重叠 |
| C.增大白鹇、猕猴所在的自然保护区的面积可增大其环境容纳量 |
| D.该种方法可用于某种大型乔木未来种群数量变化预测及性别比例的确定 |
您最近一年使用:0次
名校
4 . 20世纪60年代,迁徙到我国新疆地区的高鼻羚羊种群由于捕猎、迁徙路线被阻断、栖息地被破坏而消失。分布在哈萨克斯坦的高鼻羚羊种群由于盗猎、疾病等原因,在过去几十年时间内的种群数量变化如图所示。2015年高鼻羚羊感染病菌大量死亡,种群总数剩余25.67万,后因全球疫情影响,减少了人类干扰,到2022年种群恢复到156万,已达到“羊口巅峰”。下列关于该种群的说法正确的是( )

| A.从2015年到2022年高鼻羚羊种群数量呈现出“J”形增长 |
| B.准确统计高鼻羚羊的种群数量可以采取标记重捕法 |
| C.2015年高鼻羚羊种群数量变化的原因是密度制约因素引起的 |
| D.人类生活的干扰以及疾病是限制高鼻羚羊种群数量增长的重要因素 |
您最近一年使用:0次
真题
5 . 高寒草甸是青藏高原主要的生态系统,多年来受气候变化和生物干扰的共同影响退化严重。高原鼢鼠广泛分布于青藏高原高寒草甸,常年栖息于地下。有研究发现,高原鼢鼠挖掘洞道时形成的众多土丘,能改变丘间草地的微生境土壤物理性状,进而对该栖息生境下植物群落的多样性、空间结构以及物种组成等产生显著影响。随着高原鼢鼠干扰强度增大,鼠丘密度增加,样地内植物物种数明显增多,鼠丘间原优势种在群落中占比减少,其他杂草的占比逐渐增加。回答下列问题:
(1)调查鼠丘样地内高原鼢鼠的种群密度,常采用的方法是________ 。
(2)高原鼢鼠干扰造成微生境多样化,为栖息地植物提供了更丰富的________ ,促进植物群落物种共存。
(3)如果受到全球气候变暖加剧以及人为干扰如过度放牧等影响,高寒草甸生态系统发生逆行演替,其最终生态系统类型可能是________ 。与高寒草甸生态系统相比,演替后的最终生态系统发生的变化是________ (填序号)。
①群落结构趋于简单,物种丰富度减少 ②群落结构不变,物种丰富度增加 ③群落结构趋于复杂,物种丰富度减少 ④群落结构趋于简单,物种丰富度增加
(4)在高原鼢鼠重度干扰的地区,如果需要恢复到原有的生态系统,从食物链的角度分析,可以采用的措施是________ ,其原理是________ 。
(5)上述材料说明,除了人为活动、气候变化外,群落演替还受到________ 等生物因素的影响(回答一点即可)。
(1)调查鼠丘样地内高原鼢鼠的种群密度,常采用的方法是
(2)高原鼢鼠干扰造成微生境多样化,为栖息地植物提供了更丰富的
(3)如果受到全球气候变暖加剧以及人为干扰如过度放牧等影响,高寒草甸生态系统发生逆行演替,其最终生态系统类型可能是
①群落结构趋于简单,物种丰富度减少 ②群落结构不变,物种丰富度增加 ③群落结构趋于复杂,物种丰富度减少 ④群落结构趋于简单,物种丰富度增加
(4)在高原鼢鼠重度干扰的地区,如果需要恢复到原有的生态系统,从食物链的角度分析,可以采用的措施是
(5)上述材料说明,除了人为活动、气候变化外,群落演替还受到
您最近一年使用:0次
6 . 2023年底,“哈尔滨”周边的雪乡国家森林公园成为旅游热门地。雪乡国家森林公园始建于1999年,在政府及相关研究人员的努力下,逐渐从单一的“伐木林”向综合型的森林公园发展。
请结合材料回答下列问题:
(1)雪乡森林公园具有“夏无三日晴,冬雪漫林间”的特点,在该条件下,形成以松、杉、柳、锻树为优势种的森林群落,在调查森林植物物种数量的变化时,为使数据准确可靠,应采用的研究方法包括___(多选)。
(2)雪乡森林公园中,乔木层、灌木层和草本层的物种数量存在显著差异,且公园内不同区域的植物呈镶嵌性分布,分别体现了群落的___ 结构。
(3)对森林进行适度的采伐不会破坏生态系统的结构和功能,这是因为生态系统具有___ 稳定性。研究发现,1999年前的雪乡储碳量出现逐年下降的现象,试分析可能的原因是___ 。
(4)云杉梢斑螟是危害云杉等林木的主要害虫之一。该地区的云杉梢斑螟构成了生命系统层次中的一个___ 。可用黑灯诱捕法防治云杉梢斑螟,该方法利用了云杉梢斑螟具有___ 的特点。为了研究云杉梢斑螟的防治,技术人员要调查云杉梢斑螟的___ ,来评估防治效果。
(5)在政府及相关研究人员的努力下,植被面积和植被覆盖度明显增加,多种珍稀野生动物也随之增加,原因是___ 。森林公园在维持自然界碳平衡中发挥一定作用,要实现碳中和,除了提高森林覆盖率外,还可以采取的措施有___ (答两点)等。
请结合材料回答下列问题:
(1)雪乡森林公园具有“夏无三日晴,冬雪漫林间”的特点,在该条件下,形成以松、杉、柳、锻树为优势种的森林群落,在调查森林植物物种数量的变化时,为使数据准确可靠,应采用的研究方法包括___(多选)。
| A.对公园中的植物逐个进行调查 |
| B.选择特定季节开展调查 |
| C.在高密度区域选取多个样方调查 |
| D.随机选取多个样方调查 |
(3)对森林进行适度的采伐不会破坏生态系统的结构和功能,这是因为生态系统具有
(4)云杉梢斑螟是危害云杉等林木的主要害虫之一。该地区的云杉梢斑螟构成了生命系统层次中的一个
(5)在政府及相关研究人员的努力下,植被面积和植被覆盖度明显增加,多种珍稀野生动物也随之增加,原因是
您最近一年使用:0次
7 . 生物多样性锐减是人类面临的重大环境问题之一。回答下列问题:
(1)老虎是森林中的“王者”,常用尿液标记领地,向同类传递______ 信息。每只生活在高纬地区的东北虎需要几百或上千平方公里的领地才能维持其生存,而每只生活在低纬地区的孟加拉虎则需要几十平方公里的领地。据此分析,调查野外老虎种群密度____ (填“适宜”或“不适宜”)用标记重捕法。从能量流动角度分析,相比孟加拉虎,东北虎需要更大领地的原因可能是______ 。
(2)人类活动使森林面积快速减少,老虎所处的______ 高,能获得的能量少,导致野外的老虎“销声匿迹”。目前,我国野外的华南虎已经消失,现存仅100多只人工培育的华南虎,且都是1950年捕获的6只华南虎的后代。野化华南虎困难重重,华南虎______ 的下降会导致其对复杂环境的适应能力减弱。
(3)华南虎野外灭绝后,因______ ,野猪的数量快速上升。野猪是杂食性动物,与多种动物的_____ 存在重叠,导致激烈的种间竞争,影响了许多动物的生存。
(4)生物入侵是导致生物多样性锐减的原因之一。1859年,兔子被引入澳洲,出生后半年性成熟,妊娠期仅为30天,一次生育4~12个后代,使得兔子种群具有极高的______ ,在澳洲迅速泛滥成灾。数以亿计的兔子导致澳洲大陆______ 减少,生态系统的稳定性明显下降。为防治澳洲“兔灾”,人们引入了对兔子有致命毒性的粘液瘤病毒。除病毒自身因素外,该病毒在短期内感染大量兔子的原因是______ 。
(1)老虎是森林中的“王者”,常用尿液标记领地,向同类传递
(2)人类活动使森林面积快速减少,老虎所处的
(3)华南虎野外灭绝后,因
(4)生物入侵是导致生物多样性锐减的原因之一。1859年,兔子被引入澳洲,出生后半年性成熟,妊娠期仅为30天,一次生育4~12个后代,使得兔子种群具有极高的
您最近一年使用:0次
8 . 全球气候变暖及生产生活污水的排放对渤海湾生物种群数量和群落结构造成了严重影响,科研人员为了解渤海湾水域生态状况,对入海河口(淡水河流流入海洋的入口)的浮游生物进行了调查研究。回答下列问题:
(1)海水温度属于____________ 信息,可借助空气、水等媒介传播,这些信息传播的媒介称为_________ 。
(2)海水受到严重污染会造成赤潮现象的发生,科研上常以蓝细菌作为水体污染的指示生物,来监测水体的污染程度。采集水样后,可通过________________ 的方法并借助________________ 在显微镜下对其进行观察计数。
(3)浮游动物以浮游植物、细菌或者碎屑等为食物,它对环境变化敏感,因而也可用来监测和评价水质情况。浮游动物作为物质循环中的重要环节,其作用是:①________ ;②________ 。
(4)全球气候变暖加剧了渤海湾水域浮游植物中优势种的改变,进而影响了浮游动物种类和数量的改变,群落的这种变化属于群落的________________ 。调查发现,入海河口附近的浮游动物种类和数量低于渤海湾中部,这主要体现了群落结构在________________ 方向上的改变。入海河口附近的浮游动物种类和数量低于渤海湾中部的原因可能是________________ 。因此,可依据相关浮游生物种类和数量的变化对水质情况进行监测和评价。
(1)海水温度属于
(2)海水受到严重污染会造成赤潮现象的发生,科研上常以蓝细菌作为水体污染的指示生物,来监测水体的污染程度。采集水样后,可通过
(3)浮游动物以浮游植物、细菌或者碎屑等为食物,它对环境变化敏感,因而也可用来监测和评价水质情况。浮游动物作为物质循环中的重要环节,其作用是:①
(4)全球气候变暖加剧了渤海湾水域浮游植物中优势种的改变,进而影响了浮游动物种类和数量的改变,群落的这种变化属于群落的
您最近一年使用:0次
名校
9 . “红树”是生长在海岸潮间带(涨潮及落潮水位之间的滩涂),周期性受海水浸淹的木本植物的统称。因人类活动和互花米草入侵,广东省淇澳岛红树林面积减少至仅剩32公顷,研究人员从南亚引入了速生红树植物——无瓣海桑(Sonneratia apetala)以控制互花米草蔓延,现今淇澳岛红树林面积已恢复至600公顷以上。
(1)对不同种类的红树进行特定基因测序以揭示其亲缘关系是生物进化的______证据。
(2)下图为红树林群落中植被分布情况,列举2项可能对该群落的结构有影响的非生物因素______ 。
(4)调查淇澳岛地区人工引种的无瓣海桑种群密度的常用传统手段是______ (样方法/标记重捕法)。现可通过无人机遥感技术进行替代,无人机技术亦可调查无瓣海桑树高、胸径、生物量等指标,试写出一条无人机技术与传统调查手段相比的优点或缺点:______ (请注明优点或缺点)。
为揭示无瓣海桑控制互花米草蔓延的机制,研究人员开展实验得出了如表1、表2所示结果。
表1.不同林龄无瓣海桑林的部分生理指标
表2.无瓣海桑土壤浸出液对黑麦草发芽、生长的影响
注:互花米草植株高度一般为1~3m;林冠郁闭度为林冠在地面的投影面积占林地总面积的比例,影响光透过林冠;
黑麦草种子在发芽期对化学物质较为敏感,常用于土壤化感作用(指植物分泌化学物质进入土壤影响其他生物生长)的测定。
(5)无瓣海桑与互花米草的种间关系为:______ 。结合上述资料推测无瓣海桑控制互花米草蔓延的可能机制______ 。
(6)科学家设想用引入种无瓣海桑替代入侵物种互花米草,后再用本土红树物种代替无瓣海桑,按照该设想对下图所示的演替过程图片进行排序______ 。(用编号和箭头排序)该演替过程属于______ (初生/次生)演替。
(1)对不同种类的红树进行特定基因测序以揭示其亲缘关系是生物进化的______证据。
| A.化石 | B.分子生物学 | C.胚胎学 | D.比较解剖学 |
(2)下图为红树林群落中植被分布情况,列举2项可能对该群落的结构有影响的非生物因素
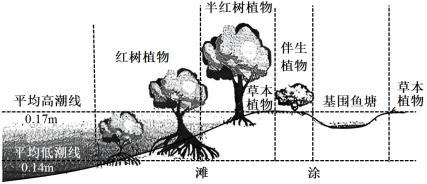
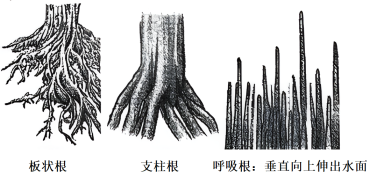
| A.板状根有利于红树承受潮汐冲击 |
| B.呼吸根有利于根部细胞获取氧气 |
| C.说明红树能主动适应潮间带环境 |
| D.是长期的变异与自然选择的结果 |
(4)调查淇澳岛地区人工引种的无瓣海桑种群密度的常用传统手段是
为揭示无瓣海桑控制互花米草蔓延的机制,研究人员开展实验得出了如表1、表2所示结果。
表1.不同林龄无瓣海桑林的部分生理指标
无瓣海桑林的林龄/a | 平均植株高度/m | 林冠层郁闭度 | 平均植株数量株/100m2 |
5 | 11.8 | 84% | 19.9 |
10 | 14.9 | 67% | 15.4 |
16 | 17.8 | 56% | 9.8 |
无瓣海桑土壤浸出液浓度(g/mL) | 黑麦草发芽率(%) | 根长(cm) | 苗高(cm) |
1.0 | 86.7 | 3.9 | 3.8 |
0.25 | 83.3 | 4.2 | 4.1 |
0.5 | 78.9 | 3.6 | 3.7 |
1.0 | 73.3 | 2.6 | 2.7 |
黑麦草种子在发芽期对化学物质较为敏感,常用于土壤化感作用(指植物分泌化学物质进入土壤影响其他生物生长)的测定。
(5)无瓣海桑与互花米草的种间关系为:
(6)科学家设想用引入种无瓣海桑替代入侵物种互花米草,后再用本土红树物种代替无瓣海桑,按照该设想对下图所示的演替过程图片进行排序
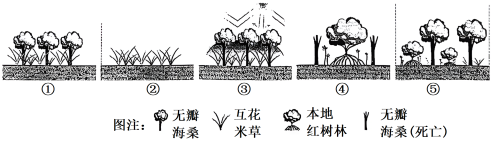
| A.无瓣海桑的扩散受海水盐度、极端气温等因素制约,扩张速度有限 |
| B.无瓣海桑在本地存在天敌,鸟类和螃蟹会捕食其种子 |
| C.无瓣海桑对部分本地红树植物生长有一定的抑制作用 |
| D.无瓣海桑在滩涂中低潮段有生存优势,而随着滩涂演替成陆地会逐渐丧失其优势 |
您最近一年使用:0次
解题方法
10 . 伞花木是我国特有的Ⅱ级重点保护的濒危野生植物。科研人员对我省龟山保护区中天然保存的伞花木群落进行调查研究,将该群落分为5个20×20m2的正方形样地,进而随机选取10×10m2的样方,对乔木层植物进行调查;另随机选取5×5m2的样方,对灌木层植物进行调查。相关结果如表1、2。请回答:
表1伞花木群落部分植物种类及数量
表2伞花木群落部分植物的生态位宽度
(注:生态位宽度是指一个物种在所处环境中能够利用资源或占据空间的范围。生态位宽度越大,物种适应环境能力和对资源的利用能力越强。)
(1)据表1结果分析,样方法除可用于调查种群密度外,还可用于调查_______ ;研究伞花木群落垂直结构时,除调查乔木层、灌木层的植物外,还应调查________ 层的植物。
(2)据表1、2结果分析,相比鸡仔木,东南栲虽然株数较多,但生态位宽度较小的原因是
_____________ 。
(3)研究伞花木的生态位时,除种群密度外,还应调查伞花木的_______ (写出2点即可)等特征。伞花木虽为濒危植物,但在本群落中的数量却不是最少,可能的原因是____________ 。
(4)以伞花木树干大小作为划分树龄标准,确定各龄级,获取伞花木种群年龄结构信息,如图。
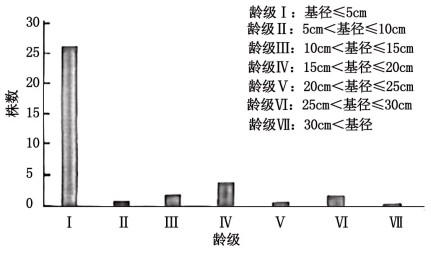
据图分析,保护伞花木应对__________ (填“幼苗”或“成株”)进行积极干预以改善其生存条件,理由是________ 。
表1伞花木群落部分植物种类及数量
层次 | 种类 | 株数 |
乔木层 | 桂花 | 120 |
青冈 | 104 | |
伞花木 | 12 | |
东南栲 | 16 | |
鸡仔木 | 8 | |
灌木层 | 桂花 | 24 |
青冈 | 18 | |
伞花木 | 26 | |
白勒 | 9 | |
臭常山 | 10 |
层次 | 种类 | 生态位宽度 |
乔木层 | 桂花 | 0.9387 |
青冈 | 0.8652 | |
伞花木 | 0.8177 | |
东南栲 | 0.3943 | |
鸡仔木 | 0.7056 | |
灌木层 | 桂花 | 0.8851 |
青冈 | 0.8444 | |
伞花木 | 0.6223 | |
白勒 | 0.4441 | |
臭常山 | 0.3614 |
(1)据表1结果分析,样方法除可用于调查种群密度外,还可用于调查
(2)据表1、2结果分析,相比鸡仔木,东南栲虽然株数较多,但生态位宽度较小的原因是
(3)研究伞花木的生态位时,除种群密度外,还应调查伞花木的
(4)以伞花木树干大小作为划分树龄标准,确定各龄级,获取伞花木种群年龄结构信息,如图。
据图分析,保护伞花木应对
您最近一年使用:0次
7日内更新
|
5次组卷
|
2卷引用:2024届福建省泉州市普通高中毕业班高三下学期5月适应性练习生物试题


