解题方法
1 . 如图所示,甲图表示有氧条件下发生在西红柿细胞内的生理反应过程,乙图表示种植某西红柿植株的密闭大棚内一昼夜空气中的CO2含量变化曲线。据图回答下列问题:________ ,能产生ATP的过程是________ (填数字)。如果光照强度突然增大,则短时间内X物质含量变化是________ 。
(2)图乙中________ (填字母)点对应的时刻光合速率等于呼吸速率,该时刻叶肉细胞中光合速率和呼吸速率的关系是________ 。经过一昼夜西红柿干重变化情况是________ 。
(3)由于植物时时刻刻进行细胞呼吸,所以一般不能一次性测量出植物的实际光合速率,某生物兴趣小组利用图丙所示的处理方法可以在光下一次性测量出实际光合速率,简述其处理的原理:________________ ,该测量方法中光合速率的表示方法是________ 。
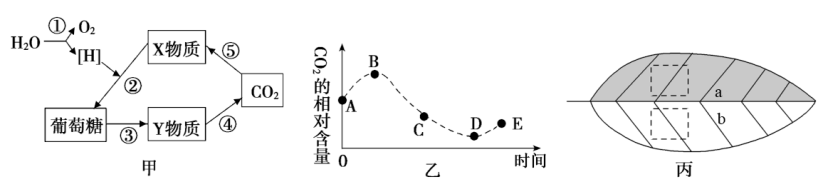
(2)图乙中
(3)由于植物时时刻刻进行细胞呼吸,所以一般不能一次性测量出植物的实际光合速率,某生物兴趣小组利用图丙所示的处理方法可以在光下一次性测量出实际光合速率,简述其处理的原理:
您最近一年使用:0次
名校
解题方法
2 . ATP合酶的作用是参与生物体的氧化磷酸化和光合磷酸化,在跨膜质子(H⁺)势能的推动下合成ATP。下列说法错误的是( )
| A.ATP合酶可位于线粒体内膜和叶绿体类囊体薄膜上 |
| B.推测线粒体内外膜间隙中的H⁺浓度大于线粒体基质中的浓度 |
| C.人进行剧烈运动时,体内的ATP水解速率大于其合成速率 |
| D.暗反应速率降低时会影响光合磷酸化的进行 |
您最近一年使用:0次
2024-05-05更新
|
226次组卷
|
2卷引用:2024届山西省朔州市怀仁市第一中学校高三下学期四模理科综合试题-高中生物
名校
解题方法
3 . 为探究冰菜在干旱和盐胁迫下的响应和适应能力,研究人员以生长状况相同的冰菜幼苗为研究对象,设置常规(CK,60%土壤含水量)、中度干旱胁迫(D,40%土壤含水量)、盐胁迫(S,60%土壤含水量+0.6%土壤含盐量)和旱盐胁迫(IS,40%土壤含水量+0.6%土壤含盐量)4个处理组,于35天后测定冰菜的生长状况和净光合速率,结果分别如表、图所示。请回答下列问题:
_______ ;盐胁迫对冰菜的影响体现在_____ ,形成对盐胁迫环境的适应,这在生产上的指导意义是______ 。
(2)研究表明,在盐胁迫下,冰菜光合作用的暗反应过程从C3途径(CO2被固定的第一个产物是C3)转变为CAM途径(白天气孔关闭,夜晚气孔打开,CO2先被固定为草酰乙酸,随后经一系列反应重新释放出CO2并参与卡尔文循环),这种转变对冰菜生长的意义是______ ,保证冰菜在盐胁迫条件下正常生长。请根据上述信息推测,在存在CAM途径的植株中,CO2被固定的时间为______ (填“白天”“夜晚”或“夜晚和白天”)。
(3)该实验需在培养过程中保持这四组实验的光照、温度等条件相同且适宜,目的是______ 。若要比较这四组实验中冰菜的有机物合成量,需进行的操作是______ 。
| 处理 | 株高(cm) | 分枝数(相对值) | 地上部鲜质量(g) | 地下部鲜质量(g) |
| CK | 2.5 | 0.33 | 15.3 | 0.5 |
| D | 2.1 | 0 | 9.2 | 0.33 |
| S | 2.6 | 1.67 | 30.1 | 0.68 |
| DS | 1.5 | 0 | 14.6 | 0.41 |
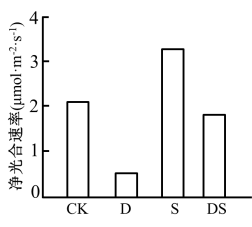
(2)研究表明,在盐胁迫下,冰菜光合作用的暗反应过程从C3途径(CO2被固定的第一个产物是C3)转变为CAM途径(白天气孔关闭,夜晚气孔打开,CO2先被固定为草酰乙酸,随后经一系列反应重新释放出CO2并参与卡尔文循环),这种转变对冰菜生长的意义是
(3)该实验需在培养过程中保持这四组实验的光照、温度等条件相同且适宜,目的是
您最近一年使用:0次
解题方法
4 . 某研究团队选择在海拔由低到高的临海、仙居、黄岩3个地区开展实验,种植适宜南方地区栽培的3个甜樱桃品种,研究不同品种的甜樱桃在不同海拔条件下的光合特性(见下表),以期为南方多山地区甜樱桃的栽培及技术推广提供参考依据。请回答下列问题:
(1)不同海拔对植物生长造成影响的环境因素主要是温度、湿度、CO2浓度和光照强度,其中影响最大的是____ ,它主要通过影响____ 的活性进而影响细胞代谢速率。
(2)光照强度主要影响光合作用的____ 阶段,此阶段发生的场所是____ 。
(3)由上表可知,在不同海拔高度条件下,甜樱桃光合特性存在显著差异,海拔越高,相对来看净光合速率越____ 。从已测参数来分析,最有可能的原因是____ 。
品种 | 地点 | 净光合速率(μmol·m-2·s-1) | 气孔导度/(mol·m-2·s-1) | 胞间CO2浓度(μL·L-1) | 果实成熟期(月/日) |
布鲁克斯 | 临海 | 2.92 | 0.0224 | 158.26 | 5/3~5/10 |
仙居 | 9.35 | 0.0744 | 167.28 | 5/10~5/20 | |
黄岩 | 13.73 | 0.1477 | 206.22 | 5/22~5/26 | |
拉宾斯 | 临海 | 4.66 | 0.0399 | 193.07 | 4/30~5/9 |
仙居 | 8.44 | 0.0664 | 164.43 | 5/7~5/13 | |
黄岩 | 13.69 | 0.1676 | 229.56 | 5/14~5/22 | |
红蜜 | 临海 | 5.88 | 0.0600 | 197.54 | 5/6~5/12 |
仙居 | 11.61 | 0.1090 | 197.54 | 5/10~5/14 | |
黄岩 | 16.88 | 0.2610 | 254.49 | 5/21~5/31 |
(2)光照强度主要影响光合作用的
(3)由上表可知,在不同海拔高度条件下,甜樱桃光合特性存在显著差异,海拔越高,相对来看净光合速率越
您最近一年使用:0次
名校
解题方法
5 . 1946年,美国科学家卡尔文等用小球藻做实验材料,将14CO2通入到盛有小球藻的玻璃器皿中,给予充足的光照(图1)。每隔一定时间取样,并将样品立即加入到煮沸的甲醇中,然后用甲醇将标记化合物提取出来,再通过双向纸层析法(图2)分离混合代谢产物,并通过一定的方法鉴定其成分。卡尔文等人根据被14C标记的化合物出现时间的先后,推测暗反应的过程。请回答下列问题:______ 。
(2)图1中灯与小球藻培养液之间盛水玻璃柱的作用是______ 。将样品立即加入到煮沸的甲醇中的目的是______ 。
(3)通过纸层析法分离混合代谢产物的原理是______ 。将层析纸晾干后,旋转90°,然后用层析液B进行第二次层析的目的是______ 。
(4)结果发现:照光30秒,14C分布于许多化合物中,缩短到1秒时,几乎所有的14C都集中在3-磷酸甘油酸上,这说明______ 。
(5)光通过光反应改变叶绿体的内部环境,间接地影响酶的活性。例如,光促进H+从叶绿体基质进入类囊体腔内,同时交换出Mg2+,使叶绿体基质中的H+和Mg2+浓度正适合与暗反应相关酶的活性。在弱光环境下,植物暗反应速率降低的原因是______ (答出两点)。

(2)图1中灯与小球藻培养液之间盛水玻璃柱的作用是
(3)通过纸层析法分离混合代谢产物的原理是
(4)结果发现:照光30秒,14C分布于许多化合物中,缩短到1秒时,几乎所有的14C都集中在3-磷酸甘油酸上,这说明
(5)光通过光反应改变叶绿体的内部环境,间接地影响酶的活性。例如,光促进H+从叶绿体基质进入类囊体腔内,同时交换出Mg2+,使叶绿体基质中的H+和Mg2+浓度正适合与暗反应相关酶的活性。在弱光环境下,植物暗反应速率降低的原因是
您最近一年使用:0次
2024-04-23更新
|
191次组卷
|
5卷引用:2024届山西省吕梁市高三下学期4月模拟考试理科综合试题-高中生物
解题方法
6 . 草莓口感独特、营养丰富,是人们喜爱的水果。为了弥补大棚草莓在秋冬季生长时光照不足的问题,科研人员以某草莓品种为材料,研究了不同颜色光照进行补光的效果,实验结果如下表所示。回答下列问题:
注:CK组的处理为不补光;T1组的处理为补红光;T2组的处理为补与T1组同等强度的白光。
(1)叶绿素含量升高后会通过增加______ (填物质)的生成速率来提高草莓的暗反应速率。测定各组叶绿素含量时,每组选取叶片时应注意________________________ ,纸层析法分离光合色素时,叶绿素a的条带在叶绿素b之上,原因是________________________ 。
(2)假设实验过程中各组实验材料呼吸作用强度不变且相同,与CK组相比,T1组的净光合速率升高,除该组植株叶片中叶绿素含量升高、固定光能增多外,其他可能原因有________________________ 。
(3)从光合色素的吸收光谱考虑,科研人员建议增设T3组,预期T3组的处理方法为____________ 。
(4)上述三组实验结果显示,在种植同样株数的草莓时,产量最高的一组是______ 。上述研究结果对大棚草莓生产的指导意义是________________________ 。
光源 | 叶绿素含量(mg·g-1) | 气孔导度(mmol·m-2·s-1) | 净光合速率(μmol·m-2·s-1) | 单果质量(g·果-1) | 结果数(个·株-1) |
CK组 | 1.13 | 308 | 16.04 | 12.11 | 9.9 |
T1组 | 2.31 | 313 | 16.32 | 14.51 | 13.2 |
T2组 | 1.79 | 310 | 15.39 | 18.07 | 12.7 |
(1)叶绿素含量升高后会通过增加
(2)假设实验过程中各组实验材料呼吸作用强度不变且相同,与CK组相比,T1组的净光合速率升高,除该组植株叶片中叶绿素含量升高、固定光能增多外,其他可能原因有
(3)从光合色素的吸收光谱考虑,科研人员建议增设T3组,预期T3组的处理方法为
(4)上述三组实验结果显示,在种植同样株数的草莓时,产量最高的一组是
您最近一年使用:0次
解题方法
7 . 为研究物质增产胺对甜瓜光合作用的影响,将生长状况相同的甜瓜幼苗均分为四组,分别测定了四组幼苗的光合速率、Rubisco(固定二氧化碳的酶)活性、丙二醛(膜脂过氧化产物,其含量与生物膜受损程度呈正相关)含量。回答下列问题:
(1)Rubisco催化CO₂固定的反应产物是________ ,发生在甜瓜叶肉细胞的________ (具体部位)。
(2)增产胺对Rubisco活性的影响在遮光条件下更大,判断依据是_________________ 。第③组光合速率最低的具体原因有____________________ (写出2点)。
(3)为研究遮光和不遮光条件下增产胺对甜瓜叶绿素含量的影响,如何继续进行实验,实验思路是:_______________ 。
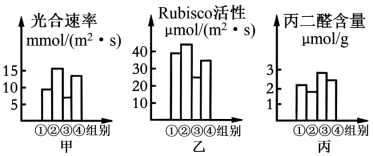
(1)Rubisco催化CO₂固定的反应产物是
(2)增产胺对Rubisco活性的影响在遮光条件下更大,判断依据是
(3)为研究遮光和不遮光条件下增产胺对甜瓜叶绿素含量的影响,如何继续进行实验,实验思路是:
您最近一年使用:0次
解题方法
8 . 图1是拟南芥进行光合作用的示意图,PSI(光系统I)和PSⅡ(光系统Ⅱ)是由蛋白质和光合色素组成的复合物。回答下列问题。__________ 。若CO2浓度降低,则图中电子传递速率会__________ (填“升高”或“降低”),原因是__________ 。
(2)强光条件下,过剩的光能会对PSⅡ复合体造成损伤,导致光合作用强度减弱。遭受强光损伤的拟南芥幼叶细胞中,叶绿素酶(CLH)基因表达量明显上升,科研人员为研究幼叶应对强光影响的机制,分别测定野生型(WT)、CLH基因缺失的突变型(clh-1)和CLH基因过量表达的突变型(clh-2)拟南芥在强光照射后的生存率,结果如图2所示。据图可知,CLH基因可以__________ 拟南芥在强光照射后的生存能力。__________ 合成受阻。
②科研人员用特定抗体检测H蛋白在叶绿体内的分布,结果如图3所示(各泳道的蛋白上样量均保持一致),依据实验结果可以得出的结论是__________ 。
③H蛋白是一种热应激蛋白(温度升高时表达),调控叶绿体基因编码的RNA聚合酶的活性。据此推测,H基因突变体在30℃时叶子呈白色的原因是__________ 。
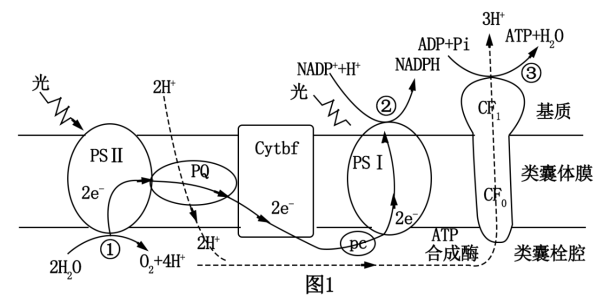
(2)强光条件下,过剩的光能会对PSⅡ复合体造成损伤,导致光合作用强度减弱。遭受强光损伤的拟南芥幼叶细胞中,叶绿素酶(CLH)基因表达量明显上升,科研人员为研究幼叶应对强光影响的机制,分别测定野生型(WT)、CLH基因缺失的突变型(clh-1)和CLH基因过量表达的突变型(clh-2)拟南芥在强光照射后的生存率,结果如图2所示。据图可知,CLH基因可以
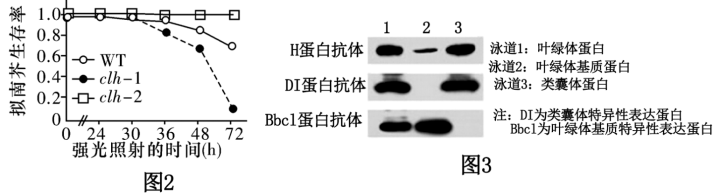
②科研人员用特定抗体检测H蛋白在叶绿体内的分布,结果如图3所示(各泳道的蛋白上样量均保持一致),依据实验结果可以得出的结论是
③H蛋白是一种热应激蛋白(温度升高时表达),调控叶绿体基因编码的RNA聚合酶的活性。据此推测,H基因突变体在30℃时叶子呈白色的原因是
您最近一年使用:0次
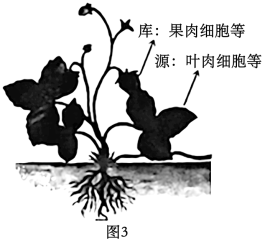
9 . 叶片是给植物其他器官提供有机物的“源”,果实是储存有机物的“库”。现以某植物为材料研究不同库源比(以果实数量与叶片数量比值表示)对叶片光合作用和光合产物分配的影响。研究发现,叶绿体中淀粉积累会导致类囊体膜结构被破坏。甲、乙、丙三组均保留枝条顶部1个果实,并分别保留大小基本一致的2、4、6片成熟叶,用13CO2供应给各组保留的叶片进行光合作用,实验结果如下表所示。请回答下列问题:
(1)该实验的自变量是______ ,该实验中采用13CO2进行光合作用的原因是____________ (答出两点)。
(2)光反应产生的______ 可以将C3转化为有机物,运输至该植物的果实。据表可知,库源比降低导致果实单果重增加,可能原因是______ 。
(3)已知卡尔文循环的中间产物三碳糖磷酸既可用于合成淀粉,还可经叶绿体膜上的磷酸转运器运至细胞质基质中参与蔗糖的合成。光合产物从“源”向“库”运输的物质形式主要是______ (填“蔗糖”“葡萄糖”或“淀粉”);若此运输过程受阻,则对叶绿体结构的影响是____________ 。根据题中研究结果可知,农业生产中,果农为提高果实品质,常采取的措施是____________ 。
项目 | 甲组 | 乙组 | 丙组 |
库源比 | 1/2 | 1/4 | 1/6 |
净光合速率/(μmol·m-2·s-1) | 9.31 | 8.99 | 8.75 |
果实中含13C光合产物/mg | 21.96 | 37.38 | 66.06 |
单果重/g | 11.81 | 12.21 | 19.59 |
(2)光反应产生的
(3)已知卡尔文循环的中间产物三碳糖磷酸既可用于合成淀粉,还可经叶绿体膜上的磷酸转运器运至细胞质基质中参与蔗糖的合成。光合产物从“源”向“库”运输的物质形式主要是
您最近一年使用:0次
名校
解题方法
10 . 科研人员利用人工气候箱,以某品种草莓为材料,研究环境因素对其叶绿素含量和净光合速率的影响,定期对叶片进行测定,结果见图1、图2。回答下列问题: ____________ ,层析后的滤纸条上色素带颜色为____________ 的是叶绿素a,该色素分布的部位进行光合作用相关反应时需要暗反应提供的原料是____________ 。
(2)由图1、2可知,相对于低温处理,在低温弱光处理下,净光合速率下降得更明显,主要是因为______________ 。12-18d之间,对照组净光合速率下降,可能的内部因素为____________ 。
(3)如图3所示,在植物体内,制造并输出有机物的组织器官被称为“源”,接纳有机物用于生长或贮藏的组织器官被称为“库”。若要验证“源”、“库”之间存在上述物质代谢关系,可以使用14CO2饲喂草莓叶片,检测_____________ 。______________ 形式运输,若该过程受阻会导致叶绿体中的淀粉积累,进一步会导致_____________ 膜结构被破坏,从而直接影响光反应的进行。
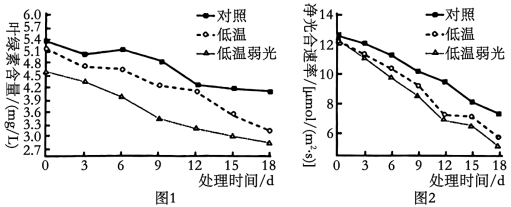
(2)由图1、2可知,相对于低温处理,在低温弱光处理下,净光合速率下降得更明显,主要是因为
(3)如图3所示,在植物体内,制造并输出有机物的组织器官被称为“源”,接纳有机物用于生长或贮藏的组织器官被称为“库”。若要验证“源”、“库”之间存在上述物质代谢关系,可以使用14CO2饲喂草莓叶片,检测
您最近一年使用:0次
2024-04-10更新
|
501次组卷
|
5卷引用:2024届山西省朔州市怀仁市第一中学校高三下学期四模理科综合试题-高中生物


