名校
1 . 依据现代生物进化理论的主要观点和内容及上述资料回答下列问题:
(1)最新群体遗传学的研究表明,现存大熊猫并未走到进化历史的尽头,仍然具有进化潜力。该种群所有个体的全部基因是这个种群的_________ 。
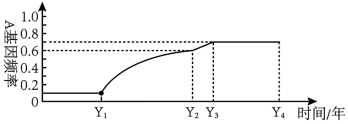
(2)在对某一种群进行调查时,发现基因型为AA和aa的个体所占的比例分别为65%和25%(各种基因型个体生存能力相同),第二年对同一种群进行的调查中,发现基因型为AA和aa的个体所占的比例分别为45%和5%,其调查的年份应该在下图的___________ 时间段内,在这一年中,该种群是否发生了进化?_________ (填“是”或“否”),理由是__________________ 。
(3)依据现代进化理论,_________ 是进化的基本单位,_________ 决定进化的方向。熊猫和其猎物因捕食关系在进化上密切相关,协同进化。协同进化是指_________ 之间、_________ 之间在相互影响中不断进化和发展。
(4)科学家经研究发现,熊猫的祖先也是食肉动物,作为捕食者,其在客观上起到了促进种群发展的作用,“收割理论”认为捕食者往往捕食_________ 的物种,这样就会避免一种或少数几种生物占绝对优势,为其他物种的形成腾出空间。捕食者的存在有利于增加_________ 。
(1)最新群体遗传学的研究表明,现存大熊猫并未走到进化历史的尽头,仍然具有进化潜力。该种群所有个体的全部基因是这个种群的
(2)在对某一种群进行调查时,发现基因型为AA和aa的个体所占的比例分别为65%和25%(各种基因型个体生存能力相同),第二年对同一种群进行的调查中,发现基因型为AA和aa的个体所占的比例分别为45%和5%,其调查的年份应该在下图的
(3)依据现代进化理论,
(4)科学家经研究发现,熊猫的祖先也是食肉动物,作为捕食者,其在客观上起到了促进种群发展的作用,“收割理论”认为捕食者往往捕食
您最近一年使用:0次
2023-08-10更新
|
196次组卷
|
2卷引用:福建省福州市连江县黄如论中学2021—2022学年高一下学期期末生物试题
名校
2 . 回答下列小题
果蝇的繁殖能力强,相对性状明显,是常用的遗传试验材料
(1)果蝇对某种杀虫剂的耐受性有两个品系:敏感型(甲)和耐受型(乙),有人做了以下两个实验。
实验一让甲品系雌蝇与乙品系雄蝇杂交,后代全为敏感型
实验二 将甲品系的卵细胞去核后,移入来自乙品系雌蝇的体细胞核,由此培育成的雌蝇再与乙品系雄蝇杂交,后代仍全为敏感型。
① 人设计实验二是为了验证___________ 。
② 另设计一个杂交实验替代实验二,该杂交实验的亲本组合为___________ 。
③如果某果蝇种群长期生活在杀虫剂浓度较高的环境中,种群数量的变化情况是___________ ,请用现代生物进化理论的观点解释这个现象出现的原因___________ 。
(2)果蝇的某一对相对性状由等位基因(N,n)控制,其中一个基因在纯合时能使合子致死(注:NN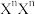 ,XnY等均视为纯合子)。有人用一对果蝇杂交,得到F1代果蝇共185只,其中雄蝇63只。
,XnY等均视为纯合子)。有人用一对果蝇杂交,得到F1代果蝇共185只,其中雄蝇63只。
①控制这一性状的基因位于___________ 染色体上,成活果蝇的基因型共有___________ 种。
②F1代雌蝇仅有一种表现型,则致死基因是___________ ,F1代雌蝇基因型为___________ 。
③F1代雌蝇共有两种表现型,则致死基因是___________ 。让F1代果蝇随机交配,理论上F2代成活个体构成的种群中基因N的频率为___________ 。
果蝇的繁殖能力强,相对性状明显,是常用的遗传试验材料
(1)果蝇对某种杀虫剂的耐受性有两个品系:敏感型(甲)和耐受型(乙),有人做了以下两个实验。
实验一让甲品系雌蝇与乙品系雄蝇杂交,后代全为敏感型
实验二 将甲品系的卵细胞去核后,移入来自乙品系雌蝇的体细胞核,由此培育成的雌蝇再与乙品系雄蝇杂交,后代仍全为敏感型。
① 人设计实验二是为了验证
② 另设计一个杂交实验替代实验二,该杂交实验的亲本组合为
③如果某果蝇种群长期生活在杀虫剂浓度较高的环境中,种群数量的变化情况是
(2)果蝇的某一对相对性状由等位基因(N,n)控制,其中一个基因在纯合时能使合子致死(注:NN
,XnY等均视为纯合子)。有人用一对果蝇杂交,得到F1代果蝇共185只,其中雄蝇63只。①控制这一性状的基因位于
②F1代雌蝇仅有一种表现型,则致死基因是
③F1代雌蝇共有两种表现型,则致死基因是
您最近一年使用:0次
3 . 碳青霉烯类抗生素是治疗重度感染的一类药物。如表为2005-2008年,该类抗生素在某医院住院患者中的人均使用量,以及从患者体内分离得到的某种细菌对该类抗生素的耐药率变化。据表回答下列问题。
(1)细菌中的耐药基因大多是因_____________ 产生的,这种细菌耐药率的变化与抗生素的使用量之间是否存在关联?_______ 依据是_____________
(2)细菌进化的实质是_____________ ,试从进化的角度解释耐药率升高的原因___________ 。
年份 | 2005 | 2006 | 2007 | 2008 |
住院患者该类抗生素的人均使用量/g | 0.074 | 0.12 | 0.14 | 0.19 |
某种细菌对该类抗生素的耐药率/% | 2.6 | 6.11 | 10.9 | 25.5 |
(1)细菌中的耐药基因大多是因
(2)细菌进化的实质是
您最近一年使用:0次
4 . 阅读科普短文,请回答问题:
尼古丁抗虫:昂贵的代价
植物和昆虫的关系可以分为两类:一类是相互受益,比如,昆虫给植物传播花粉,而植物为昆虫提供食物,可谓相得益彰,互利互惠;然而,自然界中更普遍的植物和昆虫的关系却是第二类——单纯的取食行为。在数万年的进化中植物一边需要植食性昆虫帮忙传粉,一边又要将植食性昆虫的食量控制在自己可以接受的范围,于是植物不停的进化出一系列的抗虫防御反应,而植食性昆虫也在不断适应植物的防御反应过程中进化。
尼古丁就是一种是非常好的抗虫化合物。烟草属植物被昆虫咬食后,植物的感受系统就会发出被攻击的信号,这个信号通过一套非常复杂的机制最终传到“生产工厂”,命令其生产尼古丁,然后利用这种神经毒素来进行防御。
然而植物利用尼古丁来防御昆虫是非常昂贵的,尼古丁里含有氮元素,而氮是用来合成植物生长所需要的营养成分。因此,合成抗虫用的尼古丁会降低植物的生长速率。尼古丁由植物根部生产出来后,被运送到茎叶。昆虫吃了含有尼古丁的叶片后,需要有“解毒”的过程,如果没有昆虫进化出解毒机制,就会中毒身亡。
但昆虫也会进化,进化出抵抗植物抗虫化合物的能力。如,烟草天蛾就是一种非常耐受尼古丁的昆虫。烟草天蛾在摄取了大量的尼古丁后,相应基因会被激活,使得尼古丁进入血液后不会被代谢,而是暂时储存在血液中,遇到天敌时,就将尼古丁以气态的形式通过气孔喷出。其天敌——狼蛛在闻到烟草天蛾幼虫身上浓浓的“烟味”后,会放弃对它的捕食,狼狈逃开。
尼古丁只是烟草属植物所特有的抗虫方式之一。自然界中还有很多很多相似的现象。植物为了生存,发展了这么多项技能,可是这些技能渐渐变得不像最初那么有效,未来,植物们又将运用什么手段与捕食者们斗智斗勇呢?
(1)未被昆虫捕食时,烟草属植物根部吸收的N元素可以用于细胞内合成________(多选)
(2)文中划线部分,在生物学上称之为________ 现象
(3)不同种类的昆虫通过不同方式获得抵抗植物防御反应的能力,从________ 水平为生物的趋同演化提供证据。
(4)请用达尔文的自然选择学说解释“烟草天蛾”抗尼古丁适应性的产生。
尼古丁抗虫:昂贵的代价
植物和昆虫的关系可以分为两类:一类是相互受益,比如,昆虫给植物传播花粉,而植物为昆虫提供食物,可谓相得益彰,互利互惠;然而,自然界中更普遍的植物和昆虫的关系却是第二类——单纯的取食行为。在数万年的进化中植物一边需要植食性昆虫帮忙传粉,一边又要将植食性昆虫的食量控制在自己可以接受的范围,于是植物不停的进化出一系列的抗虫防御反应,而植食性昆虫也在不断适应植物的防御反应过程中进化。
尼古丁就是一种是非常好的抗虫化合物。烟草属植物被昆虫咬食后,植物的感受系统就会发出被攻击的信号,这个信号通过一套非常复杂的机制最终传到“生产工厂”,命令其生产尼古丁,然后利用这种神经毒素来进行防御。
然而植物利用尼古丁来防御昆虫是非常昂贵的,尼古丁里含有氮元素,而氮是用来合成植物生长所需要的营养成分。因此,合成抗虫用的尼古丁会降低植物的生长速率。尼古丁由植物根部生产出来后,被运送到茎叶。昆虫吃了含有尼古丁的叶片后,需要有“解毒”的过程,如果没有昆虫进化出解毒机制,就会中毒身亡。
但昆虫也会进化,进化出抵抗植物抗虫化合物的能力。如,烟草天蛾就是一种非常耐受尼古丁的昆虫。烟草天蛾在摄取了大量的尼古丁后,相应基因会被激活,使得尼古丁进入血液后不会被代谢,而是暂时储存在血液中,遇到天敌时,就将尼古丁以气态的形式通过气孔喷出。其天敌——狼蛛在闻到烟草天蛾幼虫身上浓浓的“烟味”后,会放弃对它的捕食,狼狈逃开。
尼古丁只是烟草属植物所特有的抗虫方式之一。自然界中还有很多很多相似的现象。植物为了生存,发展了这么多项技能,可是这些技能渐渐变得不像最初那么有效,未来,植物们又将运用什么手段与捕食者们斗智斗勇呢?
(1)未被昆虫捕食时,烟草属植物根部吸收的N元素可以用于细胞内合成________(多选)
| A.淀粉 | B.尼古丁 | C.蛋白质 | D.ATP |
(3)不同种类的昆虫通过不同方式获得抵抗植物防御反应的能力,从
(4)请用达尔文的自然选择学说解释“烟草天蛾”抗尼古丁适应性的产生。
您最近一年使用:0次
5 . 在研究斑马鱼的Capm3a基因功能时,科学家发现一种遗传补偿现象:若抑制Capm3a基因的表达,斑马鱼的肝脏变小,而通过诱变使该基因转录产物中提前出现终止密码子,其转录产生的无义mRNA可以通过上调其同源基因(由同一个基因演化而来,序列相似)的表达来弥补其功能,斑马鱼的肝脏大小没有变化。具体机制如下图所示。

(1)Upf3a蛋白介导的途径可以识别同源基因的原因是__________________________ 。
(2)通过遗传补偿机制,部分无义突变的基因也被保留下来,这说明遗传补偿机制对个体生存的意义是______________ 。无义突变的保留对种群进化的意义是_____________ 。
(3)利用上述可产生无义mRNA的Capn3a基因突变体,通过诱变Upf3a基因,设计实验验证Upf3a蛋白是遗传补偿机制中的必需蛋白。
实验思路:________________________________________ 。
预期实验结果:____________________________________ 。
(1)Upf3a蛋白介导的途径可以识别同源基因的原因是
(2)通过遗传补偿机制,部分无义突变的基因也被保留下来,这说明遗传补偿机制对个体生存的意义是
(3)利用上述可产生无义mRNA的Capn3a基因突变体,通过诱变Upf3a基因,设计实验验证Upf3a蛋白是遗传补偿机制中的必需蛋白。
实验思路:
预期实验结果:
您最近一年使用:0次
6 . 大凤蝶为ZW型性别决定生物,在某种成年大凤蝶中雄蝶以宝蓝色为主,也有少量黑色;雌蝶全为黑色。大凤蝶的腹部体表形态有光滑和粗糙之分,且光滑对粗糙为显性。现有一只光滑宝蓝雄蝶和一只粗糙黑色雌蝶杂交得F1,F1具体情况如下表,请回答下列问题。(假设W染色体上不存在控制这两种性状的基因)
(1)据表推测,大凤蝶翅膀的颜色中,_______________ 为显性性状。控制该性状的基因位于_______________ 染色体上,判断依据为_____________________________________________ 。
(2)有人推测根据表中后代F1的表现型及比例,无法判断控制体表形态的基因位于常染色体上还是Z染色体上,某同学从F1,中选择光滑雌蝶和粗糙雄蝶进行一次杂交实验来判断控制体表形态的基因位置,写出预期的结果和结论。
预期的结果和结论:_____________________________________________________________ 。
(3)有些大凤蝶的幼虫蜕皮后,会变成可以融入周围草木的鲜艳的绿色,以防止天敌的捕食,从遗传水平看,这种性状出现的原因是_______________ ,从达尔文进化观点来看,这种特征的普遍形成是_______________ 的结果。
亲本 | F1 | |||||
光滑宝蓝 雄蝶×粗 糙黑色雌蝶 | 雄 | 雌 | ||||
光滑宝蓝 | 光滑黑色 | 粗糙宝蓝 | 粗糙黑色 | 光滑黑色 | 粗糙黑色 | |
612 | 209 | 597 | 195 | 807 | 823 | |
(1)据表推测,大凤蝶翅膀的颜色中,
(2)有人推测根据表中后代F1的表现型及比例,无法判断控制体表形态的基因位于常染色体上还是Z染色体上,某同学从F1,中选择光滑雌蝶和粗糙雄蝶进行一次杂交实验来判断控制体表形态的基因位置,写出预期的结果和结论。
预期的结果和结论:
(3)有些大凤蝶的幼虫蜕皮后,会变成可以融入周围草木的鲜艳的绿色,以防止天敌的捕食,从遗传水平看,这种性状出现的原因是
您最近一年使用:0次
2021-12-11更新
|
223次组卷
|
2卷引用:江西省鹰潭市2021—2022学年高二上学期期末生物试题
7 . 荠菜的果实形状有三角形和卵圆形两种,其性状的遗传涉及两对等位基因,分别用A、a,B、b表示。为探究荠菜果实形状的遗传规律,进行了杂交实验(如下图)。请回答:
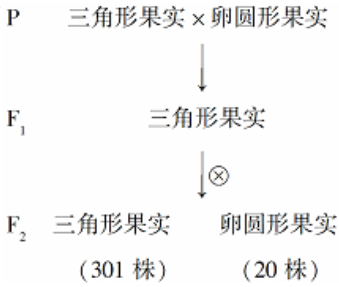
(1)图中亲本基因型分别为___________ 和_________ 。根据F2表现型比例判断,荠菜果实形状的遗传遵循__________ 定律。F1测交后代的表现型及比例为_________ 。F1测交后代的基因型及比例为________ 。
(2)另选两种基因型的亲本杂交,F1和F2的性状表现及比例与图中结果相同,推断亲本基因型为___________ 和____________ 。
(3)荠菜果实形状的相关基因a、b分别由基因A、B突变形成,基因A、B也可以突变成其他多种形式的等位基因,这体现了基因突变具有___________ 的特点。自然选择可积累适应环境的突变,使种群的基因频率_________ (填“定向”或“不定向”)改变,导致生物进化。
(1)图中亲本基因型分别为
(2)另选两种基因型的亲本杂交,F1和F2的性状表现及比例与图中结果相同,推断亲本基因型为
(3)荠菜果实形状的相关基因a、b分别由基因A、B突变形成,基因A、B也可以突变成其他多种形式的等位基因,这体现了基因突变具有
您最近一年使用:0次
2021-11-07更新
|
168次组卷
|
2卷引用:陕西省渭南市临渭区2020-2021学年高一下学期期末教学质量检测生物试题
8 . 玉米在高温、干旱条件下仍能保持高效的光合作用与其叶片的特殊结构有关。图1为玉米叶片横切图,其维管束外围的维管束鞘细胞和叶肉细胞含有不同类型的叶绿体。叶肉细胞中的叶绿体具有发达的基粒,维管束鞘细胞的叶绿体中几乎无基粒,却有很多的淀粉粒。图2为玉米光合作用的部分途径。

(1)玉米光合作用的光反应阶段在___________ 细胞中完成,暗反应阶段中C3的还原在__________ 细胞中完成。
(2)高温、干旱时植物关闭气孔,玉米在此条件下还能保持高效光合作用的原因是___________ 。玉米光合作用的途径体现了生物对环境的适应,是长期____________ 的结果。
(3)科研人员将玉米体内控制合成高活性PEP羧化酶的基因转入水稻体内,以期提高水稻在高温干旱环境下的光合速率。下表为实验测定的野生型水稻和转基因水稻的相关指标。
①与野生型水稻相比,转基因水稻______________ ,说明“转入的基因正常表达,但水稻的光合速率反而降低”。
②综合题目信息分析转基因水稻光合速率降低的原因可能是:水稻叶片的维管束鞘细胞中不含叶绿体,其光反应和暗反应均在叶肉细胞中进行。转入水稻体内的PEP羧化酶基因正常表达后,叶肉细胞中的苹果酸合成量会____________ ,但实验测定结果显示其含量降低,说明该物质_____________ ,这一过程会消耗大量NADP+,与同样发生在叶肉细胞中的_____________ 阶段竞争该物质,最终导致光合速率降低。
(1)玉米光合作用的光反应阶段在
(2)高温、干旱时植物关闭气孔,玉米在此条件下还能保持高效光合作用的原因是
(3)科研人员将玉米体内控制合成高活性PEP羧化酶的基因转入水稻体内,以期提高水稻在高温干旱环境下的光合速率。下表为实验测定的野生型水稻和转基因水稻的相关指标。
植物材料 | 叶肉细胞中苹果酸含量(μmol·g-1) | NADPH/NADP+比值 | PEP羧化酶活性(μmol·mg-1·h-1) | 放氧速率(μmol·m-2·s-1) |
野生型水稻 | 39.68 | 0.39 | 47.85 | 10.9 |
转基因水稻 | 32.96 | 2.20 | 1331.24 | 7.3 |
①与野生型水稻相比,转基因水稻
②综合题目信息分析转基因水稻光合速率降低的原因可能是:水稻叶片的维管束鞘细胞中不含叶绿体,其光反应和暗反应均在叶肉细胞中进行。转入水稻体内的PEP羧化酶基因正常表达后,叶肉细胞中的苹果酸合成量会
您最近一年使用:0次
2021-01-29更新
|
823次组卷
|
3卷引用:北京市顺义区2020-2021学年高三上学期期末生物试题(高考一模)
北京市顺义区2020-2021学年高三上学期期末生物试题(高考一模)2021届北京市顺义区高三第一次统练 生物试题(已下线)热点01 光合作用与细胞呼吸的原理与变化-2022年高考生物【热点·重点·难点】专练(北京新高考新题型)
9 . 类黄酮在植物的抗病防御反应中发挥着重要作用。苹果中类黄酮含量差异较大,新疆野生红绵肉苹果类黄酮含量高,但产量低不易储存运输,现有苹果品种产量高但类黄酮含量低。科研人员通过杂交培育类黄酮含量高且易储存运输苹果品系。
(1)新疆野生苹果的高黄酮特性是长期___________ 的结果。
(2)苹果多数自交不亲和、杂合程度高及幼苗期长等特征,通常批量繁殖采取________________ 等无性繁殖手段,获得稳定遗传的苹果品系。
(3)新疆野生红绵肉苹果中的“塔尔阿尔玛”和“夏红肉”与其他现有苹果品种杂交,F1苗期观测部分结果见下表:
在上述杂交过程中,“金帅×寒富”杂交组合的作用是_______________ ,其他组F1在第3 年的开花株率均在15%以上,说明新疆野生苹果的早开花结果的性状为_______________ (显/隐性)性状。
(4)新疆红绵肉苹果(相关基因均为杂合)与其他苹果品种杂交,产生的子代群体中相关性状所占比例如下表。
①依据表中数据判断绵肉和脆肉性状_______ (是/不是)由一对等位基因控制。
②若绵肉和脆肉性是由非同源染色体上两对等位基因(A/a、B/b)控制,参考表中数推测子代中脆肉植株基因型应为______________ 。
③若红肉和白肉性状由非同源染色体上两对等位基因(D/d、E/e)控制,参考表中数据推测子代中白肉植株基因型应为______ 。而实验结果与推测有些偏差,推测可能是环境因素的影响。
(5)F1的红脆株系中红肉面积越大,类黄酮含量越高,表明红肉性状与类黄酮含量正相关,已知控制类黄酮含量的基因由一对等位基因(F/f)控制,据此推测红肉基因与控制类黄酮含量的基因在染色体上的位置很可能是________ (请在图中注明F/f位置)。

(1)新疆野生苹果的高黄酮特性是长期
(2)苹果多数自交不亲和、杂合程度高及幼苗期长等特征,通常批量繁殖采取
(3)新疆野生红绵肉苹果中的“塔尔阿尔玛”和“夏红肉”与其他现有苹果品种杂交,F1苗期观测部分结果见下表:
亲本 | 杂种F1株数 | 开花株数(开花当年结果) | 幼苗期(年) | |||
第3年 | 第4年 | 第5年 | 第6年 | |||
塔尔阿尔玛×金帅 | 93 | 21 | 83 | 93 | — | 2.33—4.33 |
塔尔阿尔玛×新红星 | 107 | 18 | 78 | 107 | — | 2.33—4.33 |
塔尔阿尔玛×寒富 | 96 | 21 | 81 | 96 | — | 2.33—4.33 |
塔尔阿尔玛×红富士 | 110 | 17 | 82 | 110 | — | 2.33—4.33 |
夏红肉×红富士 | 126 | 19 | 92 | 126 | — | 2.33—4.33 |
金帅×寒富 | 336 | 0 | 27 | 209 | 336 | 3.33—5.33 |
在上述杂交过程中,“金帅×寒富”杂交组合的作用是
(4)新疆红绵肉苹果(相关基因均为杂合)与其他苹果品种杂交,产生的子代群体中相关性状所占比例如下表。
亲本 | 子代株数 | 子代相关性状所占比例 | ||
♀红肉绵肉 | ♂白肉脆肉 | 红肉(%) | 脆肉(%) | |
塔尔阿尔玛 | 金帅 | 93 | 33.3 | 24.0 |
塔尔阿尔玛 | 新红星 | 107 | 37.4 | 24.3 |
塔尔阿尔玛 | 寒富 | 96 | 32.3 | 26.7 |
塔尔阿尔玛 | 红富士 | 110 | 35.5 | 27.5 |
夏红肉 | 红富士 | 126 | 26.2 | 23.9 |
①依据表中数据判断绵肉和脆肉性状
②若绵肉和脆肉性是由非同源染色体上两对等位基因(A/a、B/b)控制,参考表中数推测子代中脆肉植株基因型应为
③若红肉和白肉性状由非同源染色体上两对等位基因(D/d、E/e)控制,参考表中数据推测子代中白肉植株基因型应为
(5)F1的红脆株系中红肉面积越大,类黄酮含量越高,表明红肉性状与类黄酮含量正相关,已知控制类黄酮含量的基因由一对等位基因(F/f)控制,据此推测红肉基因与控制类黄酮含量的基因在染色体上的位置很可能是
您最近一年使用:0次
10 . 螺旋蝇是家畜的毁灭性物种。在实验室里对两组数量相同的螺旋蝇进行不同的处理:一组使用杀虫剂;另一组使用电离辐射,促使雄性不育。实验结果如下图所示,请回答有关问题。
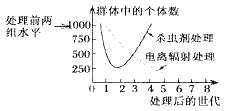
(1)用现代生物进化理论解释图中杀虫剂处理后群体中的个体数逐渐上升的原因__________________________________ 。用电离辐射促使雄性不育的方法最终能达到理想的效果,即消灭螺旋蝇,但所需时间较长,其最主要的原因是__________________ 。
(2)螺旋蝇的翅色有黄翅黑斑和橙黄黑斑两种。研究得知,黄翅黑斑(A)对橙黄黑斑(a)是显性,且亲代基因型及比例是AA(30%)、Aa(60%)、aa(10%)。若它们随机交配,则子代的基因型频率是________ 。若要使其后代的基因频率维持在这一理想状态下,除题干给出的特点外还应具备哪些条件?_______________________________ 。
(3)近年发现该种群出现了突变的白翅蝇,专家分析该种群的基因频率将会发生改变。请分析白翅基因的频率可能会怎样变化?__________________________________________ 。
(1)用现代生物进化理论解释图中杀虫剂处理后群体中的个体数逐渐上升的原因
(2)螺旋蝇的翅色有黄翅黑斑和橙黄黑斑两种。研究得知,黄翅黑斑(A)对橙黄黑斑(a)是显性,且亲代基因型及比例是AA(30%)、Aa(60%)、aa(10%)。若它们随机交配,则子代的基因型频率是
(3)近年发现该种群出现了突变的白翅蝇,专家分析该种群的基因频率将会发生改变。请分析白翅基因的频率可能会怎样变化?
您最近一年使用:0次
2019-02-19更新
|
332次组卷
|
3卷引用:【市级联考】山东省烟台市2019届高三上学期期末考试生物试题


