根据突变表型,将上述3种不同的节律突变基因分别命名为“无节律”、“长周期”和“短周期”。利用杂交获得在两条X染色体上分别携带有不同突变的雌蝇,并对其羽化节律进行测试,结果如表所示。阅读材料完成下列小题:
1.互补实验常用于确定两个突变是否影响同一个基因。以下可以说明两个隐性突变并不位于同一基因的是( )
2.请根据表中所示的数据,下列结论错误的是( )
| 基因型组成 | |||
| 第一条X染色体 | 第二条X染色体 | 测试次数 | 节律周期时长统计(hr) |
| 野生型 | 野生型 | 4 | 24.4±0.5 |
| 无节律 | 无节律 | 4 | 无节律 |
| 短周期 | 短周期 | 5 | 19.5±0.4 |
| 长周期 | 长周期 | 4 | 28.6±0.5 |
| 无节律 | 野生型 | 8 | 25.2±0.4 |
| 短周期 | 野生型 | 5 | 21.9±0.4 |
| 长周期 | 野生型 | 5 | 25.6±0.5 |
| 短周期 | 无节律 | 6 | 19.5±0.4 |
| 长周期 | 无节律 | 8 | 30.6±1.3 |
| 短周期 | 长周期 | 6 | 22.9±0.4 |
| A.杂交后代表现出正常表型 |
| B.杂交后代表现出突变特征 |
| C.杂交后代表现出中间表型 |
| D.杂交后代表型更严重 |
| A.无节律突变相对于野生型等位基因,表现为隐性突变 |
| B.即使存在野生型等位基因,短周期突变仍可影响节律 |
| C.无节律突变对短周期突变和长周期突变而言均为显性 |
| D.短周期突变对长周期突变完全显性 |
2024高二·全国·竞赛 查看更多[1]
更新时间:2024-05-22 10:03:41
|
相似题推荐
单选题-单选
|
困难
(0.15)
名校
【推荐1】野生型拟南芥的叶片是光滑形边缘,研究影响其叶片形状的基因时,发现了6个不同的隐性突变,每个隐性突变只涉及1个基因。这些突变都能使拟南芥的叶片表现为锯齿状边缘。利用上述突变培育成6个不同纯合突变体①~⑥,每个突变体只有1种隐性突变。不考虑其他突变,根据表中的杂交实验结果,下列推断正确的是( )
杂交组合 | 子代叶片边缘 |
①×② | 光滑形 |
①×③ | 锯齿状 |
①×④ | 锯齿状 |
①×⑤ | 光滑形 |
②×⑥ | 锯齿状 |
| A.导致①锯齿的突变基因与导致⑥锯齿的突变基因为等位基因 |
| B.③和④的后代与①和②的后代杂交,子代全为叶片边缘光滑形 |
| C.①和②的后代与①和⑤的后代杂交,子代叶片边缘锯齿的概率为1/2或1/4或7/16 |
| D.若②和⑤杂交,子代叶片边缘为光滑形,说明突变体⑤的突变基因与其他突变体突变基因不在同源染色体上 |
您最近一年使用:0次
单选题-题组
|
困难
(0.15)
名校
解题方法
【推荐2】阅读资料,回答下列小题:
叶绿体与线粒体间信号交流调控植物程序性细胞死亡
程序性细胞死亡(PCD)是生物体受遗传调控的自主细胞死亡现象,在植物生长发育和抵抗环境胁迫中起重要作用。植物在遭受各种生物或非生物胁迫时,体内ROS增加,ROS作为信号分子会增加线粒体膜的通透性,诱导细胞发生程序性死亡。
中国科学院遗传与发育生物学研究所李家洋研究组筛选出1个拟南芥细胞死亡突变体mod1,该突变体中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关。通过modl突变体克隆鉴定出MODI基因,该基因编码叶绿体中的脂肪酸合酶,负调控植物PCD。
近年来的研究表明,线粒体在PCD中起核心作用。一个有趣的问题是,叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD?为探明其中的作用机制,他们针对modl突变体构建了其T-DNA插入突变体库,从中筛选出能够抑制modl细胞死亡和ROS积累表型的抑制突变体,并克隆了这些抑制突变体对应的抑制基因,该基因与线粒体电子传递链复合体I(mETC复合体I)的组装及活性有关。modl是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体I的功能缺失所恢复,由此暗示植物细胞中存在叶绿体与线粒体之间的信号交流调控PCD。
最近,研究组通过大规模筛选modl突变体的抑制突变体,克隆了3个新的抑制基因pINAD-MDH、DiT l和mMDH 1。此3个基因分别编码质体定位的NAD+依赖的苹果酸脱氢酶、叶绿体被膜定位的二羧酸转运蛋白1和线粒体定位的苹果酸脱氢酶1,突变后都可抑制mod1中ROS的积累及PCD的发生。通过对这些基因进行深入的功能分析,他们论证了苹果酸从叶绿体到线粒体的转运,对线粒体中ROS的产生及随后PCD的诱导起重要作用(如图)。
1.根据上述图文信息,下列关于植物细胞中叶绿体与线粒体之间的信号交流调控PCD机制的叙述,错误的是( )
2.根据文中信息,下列证据中能够支持上述机制的是( )
叶绿体与线粒体间信号交流调控植物程序性细胞死亡
程序性细胞死亡(PCD)是生物体受遗传调控的自主细胞死亡现象,在植物生长发育和抵抗环境胁迫中起重要作用。植物在遭受各种生物或非生物胁迫时,体内ROS增加,ROS作为信号分子会增加线粒体膜的通透性,诱导细胞发生程序性死亡。
中国科学院遗传与发育生物学研究所李家洋研究组筛选出1个拟南芥细胞死亡突变体mod1,该突变体中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关。通过modl突变体克隆鉴定出MODI基因,该基因编码叶绿体中的脂肪酸合酶,负调控植物PCD。
近年来的研究表明,线粒体在PCD中起核心作用。一个有趣的问题是,叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD?为探明其中的作用机制,他们针对modl突变体构建了其T-DNA插入突变体库,从中筛选出能够抑制modl细胞死亡和ROS积累表型的抑制突变体,并克隆了这些抑制突变体对应的抑制基因,该基因与线粒体电子传递链复合体I(mETC复合体I)的组装及活性有关。modl是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体I的功能缺失所恢复,由此暗示植物细胞中存在叶绿体与线粒体之间的信号交流调控PCD。
最近,研究组通过大规模筛选modl突变体的抑制突变体,克隆了3个新的抑制基因pINAD-MDH、DiT l和mMDH 1。此3个基因分别编码质体定位的NAD+依赖的苹果酸脱氢酶、叶绿体被膜定位的二羧酸转运蛋白1和线粒体定位的苹果酸脱氢酶1,突变后都可抑制mod1中ROS的积累及PCD的发生。通过对这些基因进行深入的功能分析,他们论证了苹果酸从叶绿体到线粒体的转运,对线粒体中ROS的产生及随后PCD的诱导起重要作用(如图)。
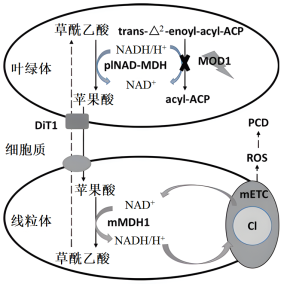
1.根据上述图文信息,下列关于植物细胞中叶绿体与线粒体之间的信号交流调控PCD机制的叙述,错误的是( )
| A.叶绿体中MOD1功能缺失导致NADH/H+在叶绿体中大量积累,草酰乙酸在pINAD-MDH作用下被氧化为苹果酸 |
| B.苹果酸通过DiT1转运到细胞质中,并进一步转移到线粒体 |
| C.在线粒体中,mMDH1将苹果酸转化为草酰乙酸,同时NADH/H+水平升高 |
| D.mETC复合体I活性增加,产生过量ROS,引发PCD |
| A.拟南芥细胞死亡突变体mod1中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关 |
| B.mod1是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体1所恢复 |
| C.抑制突变体的三个抑制基因突变后均可抑制MOD1发生PCD |
| D.苹果酸也可以由线粒体运输到叶绿体 |
您最近一年使用:0次
单选题-单选
|
困难
(0.15)
【推荐3】遗传性肾炎是一种主要表现为血尿、肾功能进行性减退、感音神经性耳聋和眼部异常的遗传性肾小球基底膜疾病,位于常染色体上的基因A4与X染色体上的基因A5等的突变会导致遗传性肾炎的发生。某遗传性肾炎家族遗传系谱图如图所示,检测了部分个体的基因型,结果如表所示。不考虑新的突变和染色体互换,下列分析正确的是( )
注1:基因A4'、基因A5'分别为基因A4、基因A5的突变基因。
注2:“+”表示有该基因,“-”表示没有该基因。

| 检测个体 | 基因A4 | 基因A4' | 基因A5 | 基因A5' |
| Ⅰ-1 | + | + | + | + |
| Ⅰ-2 | + | + | + | - |
| Ⅱ-6 | + | + | - | + |
| Ⅱ-7 | + | - | + | - |
注2:“+”表示有该基因,“-”表示没有该基因。
| A.基因A4可以突变为基因A4',说明基因突变具有随机性 |
| B.基因A4对基因A4'为隐性,基因A5对基因A5'为显性 |
| C.Ⅲ-8的基因型可能是A4A4XA5XA5'或A4A4'XA5XA5' |
| D.Ⅱ-6与Ⅱ-7生育女孩患病概率为1/2,生育男孩均不会患病 |
您最近一年使用:0次
单选题-单选
|
困难
(0.15)
【推荐1】下列技术中,无法在遗传筛选中产生新的表型的是( )
| A.EMS 诱变 | B.RNAi | C.ChIP-seq | D.CRISPR-Cas9 |
您最近一年使用:0次
单选题-题组
|
困难
(0.15)
【推荐2】有这样一个杂交:nic+×+ade,得到七种不同的子囊型。如下表所示:
(其中,nic 为烟酸依赖型,ade 为腺嘌呤依赖型)
1.nic 基因与其着丝粒之间的重组率为( )
2.nic,ade,着丝粒之间的相对位置关系是( )
子囊型 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
基因型 | +ade | ++ | ++ | + ade | + ade | ++ | ++ |
次序 | +ade | ++ | + ade | nic ade | nic + | nic ade | nic ade |
nic+ | nic ade | nic + | ++ | +ade | ++ | +ade | |
nic+ | nic ade | nic ade | nic + | nic+ | nic ade | nic+ | |
子囊数. | 808 | 1 | 90 | 5 | 90 | 1 | 5 |
(其中,nic 为烟酸依赖型,ade 为腺嘌呤依赖型)
1.nic 基因与其着丝粒之间的重组率为( )
| A.9.3% | B.5.05% | C.4.25% | D.5.2% |
| A.着丝粒-nic-ade | B.着丝粒-ade-nic | C.nic-着丝粒-ade |
您最近一年使用:0次


