解题方法
1 . 豌豆是遗传学常用的实验材料,同一植株能开很多花,不同品种植株所结种子的子叶有黄色也有绿色。现有甲、乙、丙三个品种豌豆植株,进行如表实验(相关基因用A、a表示)。下列叙述正确的是( )
实验组别 | 亲本处理办法 | 所结种子的性状及数量 | |
黄色子叶 | 绿色子叶 | ||
实验一 | 将甲植株进行自花传粉 | 409粒 | 0 |
实验二 | 将乙植株进行自花传粉 | 0 | 405粒 |
实验三 | 将甲植株的花去除未成熟的全部雄蕊,然后套上纸袋,待雌蕊成熟时,接受乙植株的花粉 | 396粒 | 0 |
实验四 | 将丙植株进行自花传粉 | 297粒 | 101粒 |
| A.实验三中,甲植株接受乙植株花粉后不需要再次进行套袋 |
| B.仅通过实验四就能推断出子叶的显隐性 |
| C.若让实验三的子代与丙植株进行杂交,理论上后代表型及比例与实验四大致相同 |
| D.取等量实验三与实验四黄色子叶种子进行混合种植,子代中约有5/12为绿色子叶种子 |
您最近一年使用:0次
2024-05-14更新
|
17次组卷
|
2卷引用:江西省宜春市2023-2024高一下学期4月质量检测生物试题
解题方法
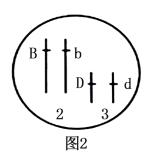
2 . 某植株(2n=16)花色有黄色、白色2种,由两对基因B、b(位于2号染色体)和D、d(位于3号染色体)共同决定,生理机制如图1。将黄花和白花植株个体杂交,F1白色:黄色=3:1,取F1中一株白色个体自交,F2中白色:黄色=13:3。回答下列问题:____________ ,两对基因通过控制____________ ,进而控制植株的花色。
(2)亲本的基因型是____________ ,F1中白花植株的基因型有____________ 种。
(3)研究发现,该植物体易被真菌感染,将抗真菌的基因E导入图2个体中,获得白花抗真菌植株,未插入的等位基因位置用e表示。每个植株的生殖力相同,子代的存活率相同。若要探究E基因插入的位置,写出实验思路并预期实验结果及结论。____________ 。
预期实验结果及结论:
①若子代的表型及其比例为白花抗真菌:黄花抗真菌:白花不抗真菌=9:3:4,则E基因与图2中B或d基因连锁;
②若子代的表型及其比例为_____________ ,则E基因与2号染色体上的b基因连锁;
③若子代的表型及其比例为_____________ ,则E基因与3号染色体上的D基因连锁;
④若子代的表型及其比例为白花抗真菌:黄花抗真菌:白花不抗真菌:黄花不抗真菌=39:9:13:3,则E基因不位于2、3号染色体上。

(2)亲本的基因型是
(3)研究发现,该植物体易被真菌感染,将抗真菌的基因E导入图2个体中,获得白花抗真菌植株,未插入的等位基因位置用e表示。每个植株的生殖力相同,子代的存活率相同。若要探究E基因插入的位置,写出实验思路并预期实验结果及结论。
预期实验结果及结论:
①若子代的表型及其比例为白花抗真菌:黄花抗真菌:白花不抗真菌=9:3:4,则E基因与图2中B或d基因连锁;
②若子代的表型及其比例为
③若子代的表型及其比例为
④若子代的表型及其比例为白花抗真菌:黄花抗真菌:白花不抗真菌:黄花不抗真菌=39:9:13:3,则E基因不位于2、3号染色体上。
您最近一年使用:0次
2024-05-07更新
|
68次组卷
|
2卷引用:江西省2024学年普通高中学业水平选择性考试生物试题
名校
解题方法
3 . 果蝇部分隐性基因在染色体上的位置如图所示。若不考虑变异,下列有关叙述正确的是( )

| A.基因lz和基因r可能出现在同一个精子中 |
| B.痕翅雄果蝇的细胞中含有4个基因r |
| C.基因dp和基因ra的遗传与性别有关联 |
| D.基因ra和基因pr为一对等位基因 |
您最近一年使用:0次
2024-05-05更新
|
52次组卷
|
2卷引用:江西省部分学校2023-2024学年高一下学期4月期中生物试题
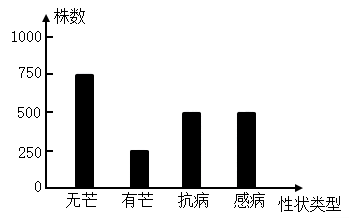
4 . 水稻是一种自花传粉植物,同时也具有异花传粉的能力,其颖片的有芒和无芒是一对相对性状,基因用A、a表示,水稻抗病与感病是另一对相对性状,基因用B、b表示,两对等位基因独立遗传。实验人员将一株无芒抗病植株和一株无芒感病植株杂交,F1的性状类型及数量如下图所示。请回答下列有关问题:_______ ,根据图中结果不能判断抗病和感病的显隐性,理由是_____ 。
(2)进一步实验发现,抗病对感病为显性,亲代无芒感病植株的基因型是______ ,子代有芒抗病植株中纯合子所占的比例为_______ 。
(3)如果将亲代无芒抗病植株进行自交得F1,F1自交,单株收获F1无芒抗病植株上的种子进行种植为一个株系,其中有______ 株系性状表型及比例为无芒抗病:无芒感病=3:1,有4/9的株系性状表型及比例为______ 。
(2)进一步实验发现,抗病对感病为显性,亲代无芒感病植株的基因型是
(3)如果将亲代无芒抗病植株进行自交得F1,F1自交,单株收获F1无芒抗病植株上的种子进行种植为一个株系,其中有
您最近一年使用:0次
解题方法
5 . 南瓜是雌雄同株异花植物,其瓜蔓丛生(A)对蔓生(a)为显性。将一株丛生南瓜与一株蔓生南瓜杂交,F1生长前期表现为丛生,生长后期表现为蔓生;F1自交,F2中丛生与蔓生在生长前期为3:1,生长后期为1:3,这种遗传现象为“蔓性转换”。下列分析正确的是( )
| A.杂交前需要对母本进行去雄、套袋处理 |
| B.F2生长前期丛生南瓜中纯合子占1/2 |
| C.发生“蔓性转换”南瓜的基因型为Aa |
| D.蔓生在南瓜生长后期属于隐性性状 |
您最近一年使用:0次
解题方法
6 . 根据以下表中实验,对三对相对性状的遗传方式推断不正确的是( )
实验序号 | 亲本组合 | 子代 |
① | 饱满豆荚豌豆(♀)×不饱满豆荚豌豆(♂) | 全为饱满豆荚豌豆 |
饱满豆荚豌豆(♂)×不饱满豆荚豌豆(♀) | 全为饱满豆荚豌豆 | |
② | 红果番茄植株甲(♀)×黄果番茄植株(乙)(♂) | 植株甲所结番茄全为红果 |
红果番茄植株甲(♂)×黄果番茄植株(乙)(♀) | 植株乙所结番茄全为黄果 | |
③ | 胚表皮深蓝色紫罗兰(♀)×胚表皮黄色紫罗兰(♂) | 全为胚表皮深蓝色 |
胚表皮深蓝色紫罗兰(♂)×胚表皮黄色紫罗兰(♀) | 全为胚表皮黄色 |
| A.实验①说明豌豆豆荚形状的遗传方式为细胞核遗传 |
| B.实验②说明番茄果皮颜色的遗传方式为细胞核遗传 |
| C.实验③说明胚表皮颜色的遗传方式为细胞质遗传 |
| D.实验②的结果不能推断番茄果实颜色的遗传方式 |
您最近一年使用:0次
7 . 某雌雄同株异花作物品系有抗虫、高产等多种优良性状,但甜度各异。为了保持其甜度,育种工作者选取甲、乙、丙三个纯合品系进行了杂交实验(若甜度由一对等位基因控制,则用A/a表示;若甜度由两对等位基因控制,则用A/a、B/b表示,依此类推),结果如表所示。回答下列问题:
(1)由表中杂交组合________ 可判断,该性状的遗传遵循自由组合定律。
(2)品系丙的基因型为________ 。若用杂交组合乙×丙的 F1微甜植株进行测交,测交后代的表型及比例为________ 。
(3)甲、乙杂交的F2中高甜植株的基因型有________ 种。若想保持这些高甜性状,则应该挑选基因型为________ 的植株用作选种,理由是________ 。
杂交组合 | F1表型 | F2表型 |
甲×乙 | 高甜 | 9/16 高甜、6/16微甜、1/16不甜 |
乙×丙 | 微甜 | 3/4 微甜、1/4 不甜 |
甲×丙 | 微甜 | 3/4 微甜、1/4不甜 |
(2)品系丙的基因型为
(3)甲、乙杂交的F2中高甜植株的基因型有
您最近一年使用:0次
2024-04-20更新
|
88次组卷
|
3卷引用:江西省上饶市世龙中学2023-2024学年高一下学期5月检测生物试卷
名校
解题方法
8 . 某观赏鱼的尾鳍分为单尾鳍、双尾鳍两种类型。研究表明,尾鳍类型受常染色体上一对等位基因(D、d)控制,雌性个体均为单尾鳍,雄性个体有单尾鳍、双尾鳍。用不同类型的此观赏鱼做了两组杂交实验,过程及结果如表所示。请据表回答
(1)由实验结果可知,控制双尾鳍性状的基因为___________ (填“显”或“隐”)性基因。此观赏鱼中,基因型为___________ 的个体只有为雄性时才能正常表达出相应性状。这对性状的遗传_____________ (填“遵循”或“不遵循”)基因的分离定律。
(2)若实验①中子代雌、雄个体随机交配,理论上其后代中双尾鳍个体所占比例为________________ 。
(3)若双尾鳍雄性与实验②中子代单尾鳍雌性杂交,所产生后代的表型和比例为__________________ 。
实验① | 实验② | |
亲代 | 单尾鳍雌性×单尾鳍雄性 | 单尾鳍雌性×双尾鳍雄性 |
子代 | 单尾鳍雌性︰单尾鳍雄子代性︰双尾鳍雄性=4︰3︰1 | 单尾鳍雌性︰单尾鳍雄性=1︰1 |
(1)由实验结果可知,控制双尾鳍性状的基因为
(2)若实验①中子代雌、雄个体随机交配,理论上其后代中双尾鳍个体所占比例为
(3)若双尾鳍雄性与实验②中子代单尾鳍雌性杂交,所产生后代的表型和比例为
您最近一年使用:0次
解题方法
9 . 玉米是雌雄同株异花的植物,玉米在自然状态下可以自交也可以杂交。玉米的甜味和非甜味受一对等位基因F/f控制。科研人员在甜味和非甜味玉米混合种植的地上收获纯合甜玉米植株上的籽粒,发现其中既有甜味籽粒也有非甜味籽粒,回答下列问题:
(1)玉米的甜味和非甜味是一对相对性状,相对性状指的是___________ 。
(2)根据题意分析,玉米的甜味和非甜味这对相对性状中,显性性状是_________ ,纯合的非甜味植株上的玉米籽粒的表型为_________ (填“全部为甜味”“全部为非甜味”或“既有甜味也有非甜味”),理由是______________ 。
(3)玉米种子的颜色有紫色和红色,受一对等位基因控制。现有紫色籽粒玉米和红色籽粒玉米若干株,试设计实验判断紫色和红色的显隐性关系,简要写出实验设计的思路____________ 。
(1)玉米的甜味和非甜味是一对相对性状,相对性状指的是
(2)根据题意分析,玉米的甜味和非甜味这对相对性状中,显性性状是
(3)玉米种子的颜色有紫色和红色,受一对等位基因控制。现有紫色籽粒玉米和红色籽粒玉米若干株,试设计实验判断紫色和红色的显隐性关系,简要写出实验设计的思路
您最近一年使用:0次
名校
解题方法
10 . 大白菜(2n)是我国重要的蔬菜作物,为两年生草本,第一年以营养生长为主,第二年春季抽薹开花。研究人员利用甲基磺酸乙酯(EMS)对大白菜萌动种子进行处理,得到早抽薹突变体甲、乙、丙,这些突变体均为单基因突变,未熟抽薹会给生产造成巨大的损失。
(1)获得该突变体采用的育种方式为____ 。研究者将早抽薹突变体甲、乙、丙分别与野生型白菜杂交,F1均为野生型,F2均出现野生型和突变体,分离比约为3:1,说明甲、乙、丙的突变性状均由____ (填“显性”或“隐性”)基因控制。
(2)SSR是DNA中的简单重复序列,非同源染色体上的SSR不同,不同品种的同源染色体上的SSR也不同。科研人员扩增出甲与野生型的若干F2代个体中的SSR序列,用于确定甲突变基因的位置,电泳结果如下图所示:____ 号染色体上,依据是____ 。
(3)为进一步判断这三个突变体所含早抽薹基因之间的位置关系,育种人员进行了杂交实验,杂交组合及F1表型见表。
(1)获得该突变体采用的育种方式为
(2)SSR是DNA中的简单重复序列,非同源染色体上的SSR不同,不同品种的同源染色体上的SSR也不同。科研人员扩增出甲与野生型的若干F2代个体中的SSR序列,用于确定甲突变基因的位置,电泳结果如下图所示:

(3)为进一步判断这三个突变体所含早抽薹基因之间的位置关系,育种人员进行了杂交实验,杂交组合及F1表型见表。
| 实验分组 | 杂交组合 | F1表型 |
| 第1组 | 甲×乙 | 早抽薹 |
| 第2组 | 甲×丙 | 野生型 |
| 第3组 | 乙×丙 | 野生型 |
实验结果表明,甲的突变基因与 ”形式表示,其中竖线表示染色体,圆点表示基因在染色体上的位置。)
”形式表示,其中竖线表示染色体,圆点表示基因在染色体上的位置。)
您最近一年使用:0次
2024-04-11更新
|
517次组卷
|
5卷引用:江西省九江市浔阳区九江第一中学2023-2024学年高三适应性考试(二模)生物试题


