名校
解题方法
1 . 某种植物是一种二倍体植物,其性别有两性株(既开雌花又开雄花)和雌株(只开雌花),由一对等位基因A、a控制;该植物的株高和叶形分别由另外两对等位基因M、m和N、n控制。研究人员将外源的DNA片段导入由纯合的高秆掌状叶两性株获得的外植体,经组织培养获得了若干株转基因植株。为探究外源DNA片段插入的位置和数量,研究人员利用获得的转基因植株进行了如下杂交实验,已知外源DNA片段不控制具体性状,但会使插入位点所在的基因突变成其等位基因。回答下列有关问题:
实验一:转基因植株甲自交,F1表现为两性株:雌株=3:1
实验二:转基因植株乙自交,F1表现为高秆掌状叶雌株:高秆掌状叶两性株:高秆柳叶两性株=1:2:1
实验三:转基因植株丙自交,F1表现为高秆掌状叶两性株:高秆柳叶两性株:矮秆掌状叶两性株:矮秆柳叶两性株=7:3:1:1。
(1)该种植物的性别表现为显性性状的是______ ;把实验一的F1均分为两组,一组自交,另一组随机交配,请问两组分别收获的F2中两性株与雌株的比例是否相同?______ 。
(2)根据实验二的结果,分析转基因植株乙中至少插入______ 个外源DNA片段,插入位置为______ 。
(3)已知实验三是由于F1的雌花产生的某种卵细胞致死导致,分析致死的卵细胞基因组成为______ 。结合三组实验现象,画出纯合的高秆掌状叶两性株的一个体细胞内这三对基因和染色体的位置关系:______ 。

(4)已知利用植物的三体植株(即某同源染色体有三条)可探究基因与染色体的关系,欲探究该植物控制叶形的基因(N、n)是否位于5号染色体上,请写出相应的实验思路。可选择的实验材料:染色体数正常的掌状叶和柳叶植株若干,5号染色体为三体的掌状叶和柳叶植株若干,所有材料均为纯合体,染色体数目异常的配子与个体均存活。
实验思路:______ 。
实验一:转基因植株甲自交,F1表现为两性株:雌株=3:1
实验二:转基因植株乙自交,F1表现为高秆掌状叶雌株:高秆掌状叶两性株:高秆柳叶两性株=1:2:1
实验三:转基因植株丙自交,F1表现为高秆掌状叶两性株:高秆柳叶两性株:矮秆掌状叶两性株:矮秆柳叶两性株=7:3:1:1。
(1)该种植物的性别表现为显性性状的是
(2)根据实验二的结果,分析转基因植株乙中至少插入
(3)已知实验三是由于F1的雌花产生的某种卵细胞致死导致,分析致死的卵细胞基因组成为
(4)已知利用植物的三体植株(即某同源染色体有三条)可探究基因与染色体的关系,欲探究该植物控制叶形的基因(N、n)是否位于5号染色体上,请写出相应的实验思路。可选择的实验材料:染色体数正常的掌状叶和柳叶植株若干,5号染色体为三体的掌状叶和柳叶植株若干,所有材料均为纯合体,染色体数目异常的配子与个体均存活。
实验思路:
您最近一年使用:0次
2023-10-21更新
|
228次组卷
|
3卷引用:湖南省株洲市一中2021届第一次模拟检测高考生物学科试题变式题16-19
2 . 玉米(2n=20)是一年生雌雄同株异花传粉的植物。现阶段我国大面积种植的玉米品种均为杂合子,杂交种(F1)的杂种优势明显,在高产、抗病等方面杂合子表现出的某些性状优于其纯合亲本(纯系)。现阶段,我国大面积推广种植的优质、高产玉米品种,均为杂合子。
(1)玉米的大穗杂种优势性状由两对等位基因(B1B2C1C2)共同控制,两对等位基因都纯合时表现为衰退的小穗性状。若大穗杂交种(B1B2C1C2)自交后代出现衰退的小穗性状的概率为1/2,则说明__________ 。如果玉米的某杂种优势性状由n对等位基因控制,且每对等位基因都独立遗传。若某杂种优势品种n对基因都杂合(亲本),其后代n对基因都纯合时才表现衰退,该品种自然状态授粉留种,F2表现衰退的概率为_________ 。
白粉病是一种造成玉米严重减产的病害。 科研人员利用基因工程技术将抗白粉病基因随机转入玉米中,获得抗病株。为进一步研究抗病株的遗传特性,让抗病株与原种进行杂交,结果如表所示:
(2)由此可知,相对于原种的感病性状,抗病性状为 _______ 性。根据F2的表型及比例能否判断控制这对性状的基因的遗传遵循分离定律还是自由组合定律?请说明理由______________________ 。
(3)玉米籽粒胚乳的颜色有黄色、紫色和杂色,科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制。已知胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。胚乳是含有一整套精子染色体的三倍体。
研究人员对胚乳颜色形成的机制作出如下推测:
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。
表2
研究发现,甲在产生配子时,10号染色体分离有时发生异常,但不影响配子的育性。表2中F1出现少量基因型为RRrr的胚乳的原因是__________ 。 表2中杂交结果仅支持推测__________ ,理由是__________ 。
(1)玉米的大穗杂种优势性状由两对等位基因(B1B2C1C2)共同控制,两对等位基因都纯合时表现为衰退的小穗性状。若大穗杂交种(B1B2C1C2)自交后代出现衰退的小穗性状的概率为1/2,则说明
白粉病是一种造成玉米严重减产的病害。 科研人员利用基因工程技术将抗白粉病基因随机转入玉米中,获得抗病株。为进一步研究抗病株的遗传特性,让抗病株与原种进行杂交,结果如表所示:
组别 | 亲本 | F1 | F2表型及比例 |
1 | ♀抗病株x ♂原种 | 自交 | 原种:抗病株≈4.93:1 |
2 | ♂抗病株x♀原种 | 自交 | 原种:抗病株≈4.85:1 |
(3)玉米籽粒胚乳的颜色有黄色、紫色和杂色,科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制。已知胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。胚乳是含有一整套精子染色体的三倍体。
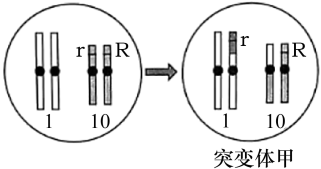
杂交组合 | F1胚乳颜色 | |
一 | 紫色RR(♀)×黄色rr(♂) | 紫色 |
二 | 紫色RR(♂)×黄色rr(♀) | 杂色 |
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。
表2
杂交组合 | 部分F1胚乳 | ||
基因型 | 颜色 | ||
三 | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
四 | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
您最近一年使用:0次
2023-06-03更新
|
300次组卷
|
3卷引用:2023年新教材山东高考生物真题变式题21-23
3 . 水稻是重要的粮食作物,如何更加有效提高水稻产量是全世界都关注的重要问题之一。
(1)在我国北方主要种植粳稻,南方主要种植籼稻。它们来自同一个祖先,由于存在________ ,导致两个 种群________ 出现差异,从而形成了两个亚种。
(2)粳稻和籼稻间的杂种优势很早就被发现,但是粳、籼杂种存在部分不育的现象。水稻的部分不育与可育是一对________ ,为探究其遗传学原理,科研人员进行了如下杂交实验。
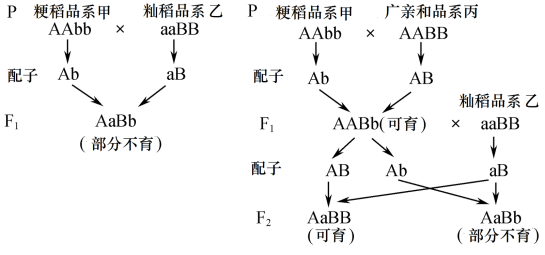
实验一:粳稻品系甲与籼稻品 系乙杂交,F1全部表现为部分不育;粳稻品系甲与广亲和品系丙杂交,F1全部表现为可育,F1与籼稻品系乙杂交,后代表现为部分不育∶可育=1∶1。研究人员根据上述现象提出一个假设,认为水稻育性由两对独立遗传的等位基因控制。具体内容如图所示:根据遗传图解推测当水稻基因组成中存在________ 基因时表现出部分不育。
(3)已知水稻的非糯性(M)和糯性(m)基因位于6号染色体上,研究人员继续进行了实验二和实验三。 ①实验二:将非糯性粳稻品系丁与糯性广亲和水稻品系己杂交,F1与非糯性籼稻品系戊杂交获得F2,结果发现F2中MM∶Mm=1∶1,且基因型为MM的个体均表现为部分不育,基因型为Mm的个体均表现为可育。F1基因型为__________ ,表现型为__________ 。
②实验三:将品系戊与品系己杂交,F1再与品系丁杂交,所得F2中基因型为MM的个体也均表现为部分不育,Mm的个体均表现为可育。根据实验二与实验三的结果 (支持,不支持)水稻的育性由两对独立遗传的等位基因控制,理由是_______________________ 。
(1)在我国北方主要种植粳稻,南方主要种植籼稻。它们来自同一个祖先,由于存在
(2)粳稻和籼稻间的杂种优势很早就被发现,但是粳、籼杂种存在部分不育的现象。水稻的部分不育与可育是一对
实验一:粳稻品系甲与籼稻品 系乙杂交,F1全部表现为部分不育;粳稻品系甲与广亲和品系丙杂交,F1全部表现为可育,F1与籼稻品系乙杂交,后代表现为部分不育∶可育=1∶1。研究人员根据上述现象提出一个假设,认为水稻育性由两对独立遗传的等位基因控制。具体内容如图所示:根据遗传图解推测当水稻基因组成中存在
(3)已知水稻的非糯性(M)和糯性(m)基因位于6号染色体上,研究人员继续进行了实验二和实验三。 ①实验二:将非糯性粳稻品系丁与糯性广亲和水稻品系己杂交,F1与非糯性籼稻品系戊杂交获得F2,结果发现F2中MM∶Mm=1∶1,且基因型为MM的个体均表现为部分不育,基因型为Mm的个体均表现为可育。F1基因型为
②实验三:将品系戊与品系己杂交,F1再与品系丁杂交,所得F2中基因型为MM的个体也均表现为部分不育,Mm的个体均表现为可育。根据实验二与实验三的结果 (支持,不支持)水稻的育性由两对独立遗传的等位基因控制,理由是
您最近一年使用:0次
名校
4 . 番茄是雌雄同株的植物,其紫茎和绿茎(由D、d控制)是一对相对性状,缺刻叶和马铃薯叶(由H、h控制)是一对相对性状,两对基因独立遗传。现利用三种不同基因型的番茄进行两组杂交,实验结果如表所示。据表分析回答下列问题。
(1)仅根据实验一的杂交的结果,能判断出__________ (填“0”或“1”或“2”)对相对性状的显隐性关系,隐性性状是___________ 。根据实验一、二的结果可知,这两对等位基因的遗传遵循__________ 定律。
(2)亲本的紫茎缺刻叶①、紫茎缺刻叶③的基因型依次是_________ 、_________ 。
(3)紫茎缺刻叶①与紫茎缺刻叶③杂交,后代的表型及比值为___________ ,后代的紫茎缺刻叶中能稳定遗传的个体占____________ 。
(4)若用实验二子代中的紫茎缺刻叶和绿茎缺刻叶植株杂交,其后代中绿茎缺刻叶植株所占的比例为_________ 。
(5)若番茄的果实颜色由两对等位基因(A和a、B和b)控制,且基因的表达与性状的关系如图1所示,为探究这两对等位基因是否位于同一对同源染色体上,某生设计了如下实验:
实验预测及结论:
①若子代晋茄果实的颜色及比例为__________ ,则A、a和B、b基因分别在两对同源染色体上。
②若子代番茄果实的颜色及比例为__________ ,则A、a和B、b基因在一对同源染色体上,且A和B在一条染色体上。
③若子代番茄果实的颜色及比例为___________ ,则A、a和B、b基因在一对同源染色体上,且A和b在一条染色体。
实验编号 | 亲本表型 | 子代表型及比例 |
实验一 | 紫茎缺刻叶①×绿茎缺刻叶② | 紫茎缺刻叶:紫茎马铃薯叶=3:1 |
实验二 | 紫茎缺刻叶③×绿茎缺刻叶② | 紫茎缺刻叶:紫茎马铃薯叶:绿茎缺刻叶;绿茎马铃薯叶=3:1:3:1 |
(1)仅根据实验一的杂交的结果,能判断出
(2)亲本的紫茎缺刻叶①、紫茎缺刻叶③的基因型依次是
(3)紫茎缺刻叶①与紫茎缺刻叶③杂交,后代的表型及比值为
(4)若用实验二子代中的紫茎缺刻叶和绿茎缺刻叶植株杂交,其后代中绿茎缺刻叶植株所占的比例为
(5)若番茄的果实颜色由两对等位基因(A和a、B和b)控制,且基因的表达与性状的关系如图1所示,为探究这两对等位基因是否位于同一对同源染色体上,某生设计了如下实验:
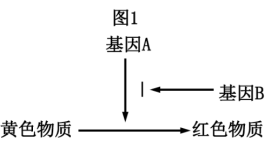
实验预测及结论:
①若子代晋茄果实的颜色及比例为
②若子代番茄果实的颜色及比例为
③若子代番茄果实的颜色及比例为
您最近一年使用:0次
2024-04-27更新
|
376次组卷
|
3卷引用:非选择题必刷题30道-备战2023-2024学年高一生物下学期期末真题分类汇编(山东专用)
5 . 水稻中存在雄性不育系。雄性可育(R)对雄性不育(r)为显性,该对基因存在于细胞核中;雄性可育(N)对雄性不育(S)存在于细胞质中,只有细胞质、细胞核均为雄性不育基因时,水稻个体才表现为雄性不育。科研人员将控制水稻大籽粒中淀粉合成的基因Qt导入雄性不育品系获得大粒正常秆品系A、大粒高秆品系B,经筛选鉴定A、B品系仅一条染色体上插入一个Qt基因。为研究基因的插入位点及对受体植株产生的影响,科研人员利用品系A、B、基因型为S(Rr)的正常粒正常秆雄性可育个体H做了以下实验,请回答下列问题:

(1)水稻的雄性不育性状是由细胞核、细胞质基因共同决定的,其中 N、R基因是否符合自由组合定律____ (填“符合”或“不符合”)?因为自由组合定律的适用条件有___________ 。水稻可育的基因型有_____ 种。
(2)实验一中,F1不可育的基因型为__________ 。选取F1中的大粒可育个体自交,F2中重粒不可育∶大粒可育∶正常粒可育比例为1∶2∶1,可推测导入的Qt基因与r基因所在染色体的位置关系______ (填“连锁”或“独立遗传”)。实验一的F2中出现了重粒性状,请据题推测其原因___________ 。(重粒是指质量重于大粒)
(3)将实验二中的大粒高秆可育水稻自交,F2个体中大粒高秆可育∶大粒高秆不育∶正常粒可育∶正常粒不育的比例为3∶1∶3∶1,而不是正常的9∶3∶3∶1,经推测出现该现象的原因是Qt基因的导入不仅使水稻植株高度发生了变化,还导致实验二的亲本所产生的某一类配子的育性降低,请分析得到比例的原因是___________ 。此时,在F2的大粒个体中,r的基因频率为____________ 。
(4)若(3)中上述推测正确,F2中大粒高秆不育个体与正常粒可育个体杂交,子代高秆可育个体所占比例为_____ 。
(1)水稻的雄性不育性状是由细胞核、细胞质基因共同决定的,其中 N、R基因是否符合自由组合定律
(2)实验一中,F1不可育的基因型为
(3)将实验二中的大粒高秆可育水稻自交,F2个体中大粒高秆可育∶大粒高秆不育∶正常粒可育∶正常粒不育的比例为3∶1∶3∶1,而不是正常的9∶3∶3∶1,经推测出现该现象的原因是Qt基因的导入不仅使水稻植株高度发生了变化,还导致实验二的亲本所产生的某一类配子的育性降低,请分析得到比例的原因是
(4)若(3)中上述推测正确,F2中大粒高秆不育个体与正常粒可育个体杂交,子代高秆可育个体所占比例为
您最近一年使用:0次
2023-02-23更新
|
573次组卷
|
3卷引用:湖南省株洲市一中2021届第一次模拟检测高考生物学科试题变式题16-19
名校
解题方法
6 . 某种昆虫为雌雄异体,其体色中的白色和灰色为一对相对性状,由A、a基因控制,其长触角与短触角为一对相对性状,与该对相对性状有关的基因为B、b,这两对等位基因的分离和组合互不干扰。科研人员进行了如下实验,
实验一:选取表型为白色和灰色的纯合亲本进行正反交,F1中出现两种结果,即全部为灰色;雄性个体为灰色,雌性个体为白色。
实验二:选取长触角(只含有B基因的纯合体)和短触角(只含有b基因的纯合体)的个体作亲本进行正反交,F1中雄性个体都表现为长触角,雌性个体都表现为短触角。
将实验二正反交得到的F1各自相互交配得到各自的F2,结果如表所示。实验过程中所获取的样本足量,不存在突变和致死现象,雌雄个体比例为1:1.下列说法正确的是( )
实验一:选取表型为白色和灰色的纯合亲本进行正反交,F1中出现两种结果,即全部为灰色;雄性个体为灰色,雌性个体为白色。
实验二:选取长触角(只含有B基因的纯合体)和短触角(只含有b基因的纯合体)的个体作亲本进行正反交,F1中雄性个体都表现为长触角,雌性个体都表现为短触角。
将实验二正反交得到的F1各自相互交配得到各自的F2,结果如表所示。实验过程中所获取的样本足量,不存在突变和致死现象,雌雄个体比例为1:1.下列说法正确的是( )
| 性别 | F2中表型及比例 |
| 雄性 | 长触角:短触角=3:1 |
| 雌性 | 长触角:短触角=1:3 |
| A.由实验一可判断该昆虫体色性状中显性性状是灰色 |
| B.由实验一可判断该昆虫的性别决定方式为XY型 |
| C.控制触角长短的基因B/b位于性染色体上 |
| D.若让F2中短触角个体相互交配,则其子代中长触角的个体所占的比例为1/6 |
您最近一年使用:0次
2024-06-04更新
|
72次组卷
|
2卷引用:2024届山东省实验中学高三下学期模拟考试生物试题
解题方法
7 . 番茄叶霉病会引起叶片发黄卷曲,导致番茄减产。为预防抗叶霉病香茄品种甲的抗病能力减弱,科研人员利用硫酸二乙酯诱变易感病番茄,获得新的抗病品种乙。科研人员利用甲、乙两个品种进行了如下杂交实验,对番茄抗叶摇病抗性进行了研究,已知甲、乙品种番茄抗叶摇病均为单基因突变引起,且甲品种抗病基因位于9号染色体上,不考虑染色体交换。请回答下列问题

(1)人工诱变育种的优点是________ 。
(2)乙品种抗叶霉病性状对易感病为________ (填“显性”或“隐性”),原因是________ .
(3)根据实验三F₁、F₂的表型及比例判断甲、乙两品种抗性基因的位置关系。
①若 F₁、F₂ 均表现为抗叶霉病,则可判断甲、乙两品种抗叶霉病基因的位置关系为________ 。
②若F₁ 表现为抗叶霉病,F₂ 中抗叶霉病个体的比例为________ ,则可判断甲、乙两品种抗叶霉病性状由位于非同源染色体上的两对等位基因控制。
(4)SSR 是 DNA 中的简单重复序列,非同源染色体上的 SSR不同,不同品种的同源染色体上的 SSR 也不同,常用于染色体特异性标记。科研人员利用PCR 技术将实验二中部分抗病个体(该部分个体基因组成及比例与 F₂理论上的基因组成及比例相同)9号染色体的 SSR进行扩增后,电泳结果如下图:

依据图中结果判断,乙品系抗叶霉病基因与9号染色体的位置关系是_________ ,理由是_______ 。
(1)人工诱变育种的优点是
(2)乙品种抗叶霉病性状对易感病为
(3)根据实验三F₁、F₂的表型及比例判断甲、乙两品种抗性基因的位置关系。
①若 F₁、F₂ 均表现为抗叶霉病,则可判断甲、乙两品种抗叶霉病基因的位置关系为
②若F₁ 表现为抗叶霉病,F
(4)SSR 是 DNA 中的简单重复序列,非同源染色体上的 SSR不同,不同品种的同源染色体上的 SSR 也不同,常用于染色体特异性标记。科研人员利用PCR 技术将实验二中部分抗病个体(该部分个体基因组成及比例与 F₂理论上的基因组成及比例相同)9号染色体的 SSR进行扩增后,电泳结果如下图:
依据图中结果判断,乙品系抗叶霉病基因与9号染色体的位置关系是
您最近一年使用:0次
名校
8 . 人类的ABO血型由IA、IB和i三个复等位基因决定,其中IA、IB基因分别决定红细胞膜上的A、B抗原,各血型及对应基因型、红细胞上的抗原、血清中抗体的关系如下表。回答下列问题:
(1)输血时,判断可以输血的标准是受血者血清中的天然抗体不会对献血者红细胞膜上的抗原发生反应。理论上AB型受血者可接受全部四种血型,理由是______________ 。
(2)一个A型血的女性的父亲是О型血,一个B型血的男性的母亲也是O型血,若该两人结婚,理论上其孩子的血型表型及比例为_______________ 。
(3)已知A、B抗原来自于前体蛋白H,而前体蛋白H由位于常染色体上的显性基因H控制,且与IA、IB、i不在同一对染色体上,若个体中只有h基因,则无法将前体物质转化为前体蛋白H,从而表现出一种稀有的孟买型血型。在一个孟买型血型的家系中,表现为O型血的个体可能的基因型有______ 种。下图为该家系的部分遗传系谱图,其中血型一定为孟买型的个体是______ ,且基因型为______ 。理论上个体1与个体2的子代表型及比例为__________________ 。
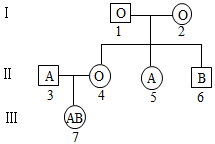
血型 | 基因型 | 红细胞上的抗原 | 血清中天然的抗体 |
A | IAI IAi | A抗原 | 抗B抗体 |
B | IBIB IBi | B抗原 | 抗A抗体 |
AB | IAIB | A抗原、B抗原 | 无 |
O | ii | 无 | 抗A抗体、抗B抗体 |
(1)输血时,判断可以输血的标准是受血者血清中的天然抗体不会对献血者红细胞膜上的抗原发生反应。理论上AB型受血者可接受全部四种血型,理由是
(2)一个A型血的女性的父亲是О型血,一个B型血的男性的母亲也是O型血,若该两人结婚,理论上其孩子的血型表型及比例为
(3)已知A、B抗原来自于前体蛋白H,而前体蛋白H由位于常染色体上的显性基因H控制,且与IA、IB、i不在同一对染色体上,若个体中只有h基因,则无法将前体物质转化为前体蛋白H,从而表现出一种稀有的孟买型血型。在一个孟买型血型的家系中,表现为O型血的个体可能的基因型有
您最近一年使用:0次
2022-10-29更新
|
339次组卷
|
4卷引用:湖南省株洲市一中2021届第一次模拟检测高考生物学科试题变式题16-19
名校
9 . 籼稻(染色体组成表示为SS)和粳稻(染色体组成表示为JJ)的杂交种具有抗逆性强、产量高的优势,但杂交种的部分花粉败育,经检测,其所有花粉中均存在某种杀死花粉的毒素蛋白,但只有育性正常的花粉中存在一种对应的解毒蛋白:进一步研究发现,编码这两种蛋白的基因均为位于籼稻12号染色体上的R区,如下图(该区的基因不发生交换)。科研人员将纯合籼稻和纯合粳稻杂交,获得F1,F1自交获得F2,F2中仅有籼—粳杂交种和籼稻,且二者比例接近1:1。

(1)F1植株产生的染色体组成为__________ 的花粉败育。若让F1作父本,籼稻作母本进行杂交,子代的染色体组成为_________ 。
(2)科研人员利用基因编辑技术分别敲除了F1中的R区相关基因,得到1~10号植株,检测其花粉育性情况,结果如下表。
(注:“++”表示全部花粉有育性:“+”表示一半花粉有育性;空白表示全部花粉败育。)
由表中结果可推测基因B和C的功能,其中编码毒素蛋白的是基因__________ 。
(3)为进一步验证基因B的作用,将基因B导入到F1中,获得转入单个基因B的F1。预期F1自交后代的染色体组成及比例是__________ (写出所有可能的比例)。
(4)请结合上述机制,推测基因C的基因频率趋于__________ (填“升高”、“降低”或“不变”),理由是__________ 。
(1)F1植株产生的染色体组成为
(2)科研人员利用基因编辑技术分别敲除了F1中的R区相关基因,得到1~10号植株,检测其花粉育性情况,结果如下表。
| F1植株编号 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| 敲除基因 | A | B | C | D | A+B | A+C | A+D | B+C | B+D | C+D |
| 花粉育性 | + | ++ | + | ++ | + | ++ | ++ |
由表中结果可推测基因B和C的功能,其中编码毒素蛋白的是基因
(3)为进一步验证基因B的作用,将基因B导入到F1中,获得转入单个基因B的F1。预期F1自交后代的染色体组成及比例是
(4)请结合上述机制,推测基因C的基因频率趋于
您最近一年使用:0次
2024-03-01更新
|
678次组卷
|
3卷引用:山东省聊城市2024年高考模拟试题(一)生物试题变式题24-25
解题方法
10 . 亲缘关系较远的水稻品系间杂交,子代具有杂种优势,有利于农业生产,但有时杂种一代会出现雄性育性下降的情况。研究发现,水稻的雄性育性与12号染色体的一段区域有关。对南方野生稻(j品系)该段染色体测序,发现该区段从上到下依次为A、B、C、D、E五个基因。对亚洲栽培稻(x品系)该段染色体测序;发现该区段只有A、B、C三个基因,基因顺序与j品系相同。为寻找影响雄性育性的基因,对j品系进行基因敲除,获得敲除纯合子(无法获得E基因敲除的纯合个体),进行系列杂交实验,结果如下表。
进一步研究发现,表中所有杂交组合的F1均能正常完成减数分裂,且杂交组合1~4的F1产生的可育花粉的12号染色体都来自j品系,不育花粉的12号染色体都来自x品系。已知杂交组合1~4的F1的花粉母细胞中D基因编码的蛋白质(减数分裂前合成,可通过减数分裂进入花粉细胞)可以导致不含E基因的花粉败育。不考虑基因突变、染色体变异和互换,回答下列问题:
(1)推测E基因的作用是___________ 。杂交组合1的F1产生的不育花粉的基因型是__________ 。杂交组合1的F1进行自交,F2产生的花粉的育性及比例是______________ 。
(2)为验证E基因的作用,若向杂交组合1的F1的花粉母细胞中来自x品系的12号染色体上转入一个E基因,则其产生的花粉__________ (填“都不育”“50%可育”或“100%可育”);若将杂交组合1的F1的花粉母细胞的E基因敲除,则其产生的花粉___________ (填“都不育”“50%可育”或“100%可育”)。
杂交组合 | 母本 | 父本 | F1花粉育性 |
1 | 纯合j品系 | 纯合x品系 | 50%的花粉可育 |
2 | A基因敲除的纯合j品系 | 纯合x品系 | 50%的花粉可育 |
3 | B基因敲除的纯合j品系 | 纯合x品系 | 50%的花粉可育 |
4 | C基因敲除的纯合j品系 | 纯合x品系 | 50%的花粉可育 |
5 | D基因敲除的纯合j品系 | 纯合x品系 | 100%的花粉可育 |
(1)推测E基因的作用是
(2)为验证E基因的作用,若向杂交组合1的F1的花粉母细胞中来自x品系的12号染色体上转入一个E基因,则其产生的花粉
您最近一年使用:0次
2024-03-09更新
|
219次组卷
|
4卷引用:山东省聊城市2024年高考模拟试题(一)生物试题变式题24-25


