检测结果 实验处理 | 净光合速率(μmol·m⁻²·s⁻¹) | |||||
品种甲 | 品种乙 | |||||
55天 | 65天 | 75天 | 55天 | 65天 | 75天 | |
对照组(常态大气浓度) | 33.0 | 33.0 | 33.0 | 30.2 | 30.2 | 30.2 |
实验组一(高CO₂浓度) | 35.3 | 35.3 | 35.3 | 33.3 | 3.43 | 3.4 |
实验组二(高O₃浓度) | 31.2 | 31.2 | 27.5 | 26.0 | 26.0 | 21.2 |
实验组三(高CO₂浓度+高O₃浓度) | 33.0 | 33.0 | 30.1 | 31.2 | 30.0 | 25.2 |
回答下列问题:
(1)光合作用中CO₂在叶绿体基质中经过
(2)若要进一步获取甲、乙两个品种水稻植株的光合速率,还要检测这两个品种水稻植株的呼吸速率,测定光合速率的思路是
(3)据表分析,长时间高浓度的O₃对水稻光合作用产生明显抑制,对
(4)理论上,大气中CO₂浓度不断升高会使绿色植物的光合作用逐渐增强。但是,有科学家认为,大气中CO₂升高带来的气温升高会使光合作用受到一定程度的抑制。请根据所学知识,从光合作用角度说明提出这个观点的理由可能是
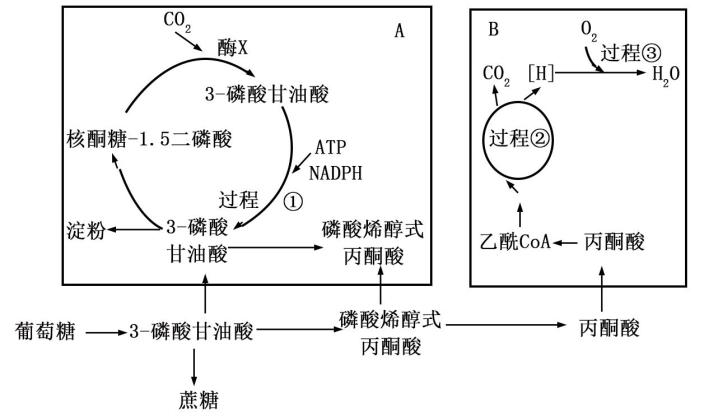
(2)在适宜光照条件下,过程③所需O2来自
(3)据图推测,大麦叶肉细胞中合成淀粉所需的酶主要存在于
(4)研究发现,大麦的光合作用和呼吸作用之间存在着紧密的联系:呼吸作用的运行能够优化光合作用。有实验者利用大麦叶片进行了下表所示实验,表中的 KCN 和 SHAM是两类呼吸抑制剂。
| 组别 | 1 | 2。 | 3 | 4 |
| 缓冲液中 的添加物 | 不添加 | 0.5 mmol/L KCN | 1mmol/L SHAM | 1mmol/L SHAM +0.5 mmol/L KCN |
| 步骤一 | 在相同位置选取大小一致且完全伸展的叶片,置于缓冲液中进行真空渗透1小时 | |||
| 步骤二 | 渗透结束后,将叶片从缓冲液中取出,清洗并吸去叶片表面残余的缓冲液; 将叶柄浸入上述缓冲液中,并将叶片置于持续的适宜光照下 | |||
| 步骤三 | 光照4小时后,使用相关仪器测定叶片的光合作用及呼吸作用速率 | |||
②该实验的目的是
镉浓度 (mg·L-1) | 气孔导度 (mmolCO2·m2·s-1) | 胞间CO2浓度 (μLm-2·s-1) | 净光合速率 (μmolCO2·m2·s-1) |
0 | 154.75 | 256.50 | 11.05 |
0.01 | 133.50 | 264.50 | 9.07 |
0.1 | 141.50 | 236.75 | 12.02 |
1 | 121.00 | 277.00 | 8.37 |
10 | 93.75 | 355.00 | 3.52 |
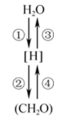
(1)①过程的场所是
(2)若该小麦长期缺Mg,则提取该小麦的色素经
(3)④过程产生的[H]来源是
(4)据表一分析,高剂量(≥1mg·L-1)镉会使气孔导度明显下降,而胞间二氧化碳浓度却增大,其主要原因是
(5)表二取阳生和阴生两种长势相似的植物,分别放在两个相同的密闭透明玻璃瓶中,在适宜温度条件下,逐渐增加光照强度,测定放氧速率的数据如下表。请回答相关问题:
光强μmol光 子/(100g·s) | 0 | 10 | 30 | 50 | 100 | 200 | 450 | 500 | |
放氧速率μ molO2/(100g·s) | 植物A | -20 | -10 | -5 | -1 | 5 | 15 | 29 | 28 |
植物B | -2 | -0.5 | 1.5 | 3 | 6 | 10 | 10 | 10 |
①光强由200μmol光子/(100g·s)突然变为450μmol光子/(100g·s)时,阳生植物短期体内的三碳酸和RuBP变化分别为
②该实验的目的是探究
③光饱和点时,阴生植物的呼吸熵为0.8(呼吸作用CO2的释放速率/呼吸作用O2的消耗速率),则该植物要从外界吸收CO2的速率是
实验一:以野生型番茄植株为实验材料进行探究,在实验第3天时测定相关实验数据,如表所示(R酶参与碳反应)
| 组别 | 温度 | 气孔导度 (mol·m²·s¹) | O₂释放速率(μmol·m²·s¹) | 胞间CO₂浓度(μmol·mol-¹) | R酶活性 (U·ml-¹) |
| 甲 | 25℃ | 99.2 | 11.8 | 282 | 172 |
| 乙 | 40℃ | 30.8 | 1.1 | 403 | 51 |
(1)光合复合体Ⅱ位于叶绿体的
(2)叶肉细胞吸收的CO₂,在
(3)由表中数据可知,CO₂浓度
(4)高温胁迫下番茄植株光反应速率减慢,分析其原因可能是:
①
②
实验二:已知叶绿素酶(CLH)能促进被破坏的D1降解。研究者以野生型番茄植株和CLH基因缺失的突变体植株为实验材料进行相关实验,测得实验结果如下图曲线所示(注:第1天之前为25℃,第1~3天为40℃,第4~5天为25℃)。
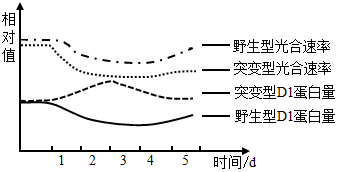
(6)研究者补充设计以
(1)光合作用过程中,CO2在
(2)用多个番茄大棚模拟不同空气污染情况对大棚温度、光合有效辐射(被光合色素吸收并转化的太阳能)相对值和光合速率相对值的影响,结果如下表。据表分析,当空气质量为重度污染时,与中度污染相比,应采取
空气质量 | 棚温/℃ | 光合有效辐射相对值 | 光合速率相对值 |
良 | 25.6 | 987 | 20.4 |
轻度污染 | 23.4 | 746 | 19.6 |
中度污染 | 23.5 | 477 | 17.1 |
重度污染 | 23.1 | 325 | 11.8 |
(3)在另一空气质量、温度、光照强度等栽培条件均适宜的番茄大棚中,将CO2浓度由375µmol·mol-1增加到750µmol·mol-1,发现光合速率并没有倍增,原因可能是
(4)给予某植物48小时持续光照,测定叶肉细胞中的淀粉积累量,结果如图1所示。为了解释该实验现象,研究人员提出了两种假设。

假设一:当叶肉细胞内淀粉含量达到一定值后,淀粉的合成停止。
假设二:当叶肉细胞内淀粉含量达到一定值后,淀粉的合成与降解同时存在。
科研人员测定了叶肉细胞的CO2吸收量和淀粉降解产物麦芽糖的含量,结果如图2所示。实验结果支持假设
为进一步确定该假设成立,研究人员在第12小时测得叶肉细胞中的淀粉含量为a,为叶片通入仅含13C标记的CO24小时,在第16小时测得叶肉细胞中淀粉总量为b,13C标记的淀粉含量为c。若a、b、c的关系满足
实验组 测量指标 | 对照 | T1 | T2 | T3 |
| 单株总干重(g) | 7.02 | 9.01 | 7.75 | 6.15 |
| 叶绿素(SPAD) | 41.52 | 46.49 | 43.84 | 35.03 |
| 叶片可溶性糖(mg/g) | 25.09 | 20.41 | 15.41 | 11.14 |
(1)绿色植物中的叶绿素主要存在于叶绿体的
(2)经过遮荫处理后,随着光照强度由强到弱,叶片可溶性糖含量逐渐降低,其原因是
(3)对照的叶片可溶性糖含量比T1,T2高,但单株总干重比T1、T2低,其原因是
(4)为验证T3的叶绿素含量降低,需要分析莲叶桐遮荫处理后光合色素含量的变化。现有干燥的定性滤纸条(其一端剪去两角,并在这一端底部1cm处画有一条细的横线)、天平、玻璃漏斗、尼龙布等充足的实验材料和用具,按步骤简要写出实验过程(根据需要可自行加步骤,不限于3步):
①
②将材料剪碎,分别置于不同研钵中,加入等量溶剂后充分研磨
③
组别 | 温度/℃ | 光照强度/(μmol·m-2·s-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/ppm | Rubisco活性/(U·mL-1) |
CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
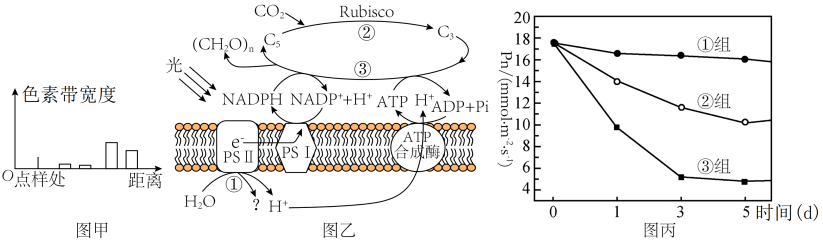
(1)分析图甲所示实验结果可知,含量最多的色素为
(2)PSII中的色素吸收光能后,将H2O分解为H+和
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降的原因
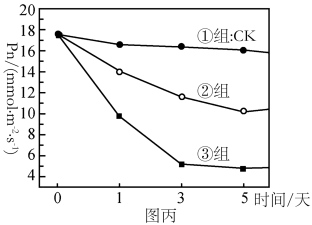
(4)D1蛋白是PSII复合物的组成部分,对维持PSII的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。已知药物SM可抑制D1蛋白的合成。为研究植物应对高温高光逆境时D1蛋白的变化机制,研究者利用番茄植株进行如下三组实验:①组在适宜温度、适宜光照下培养;③组在高温高光下培养并施加适量Sm(抑制D1蛋白合成的药物)。②组的处理方式是
材料一:某班学生进行新鲜番茄植株叶片色素的提取和分离实验,研磨时未加入CaCO3,实验结果如图甲所示。图乙是番茄植株进行光合作用的示意图,其中PSⅡ和PSⅠ是吸收、传递、转化光能的光系统。请回答下列问题:
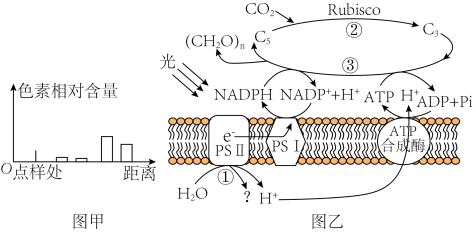
(1)分析图甲所示实验结果可知,距离点样处距离最远的色素为
(2)PSⅡ中的色素吸收光能后,将H2O分解为H+和
材料二:为研究亚高温高光对番茄光合作用的影响,某研究者将番茄植株放在CK条件(适宜温度和适宜光照)和HH条件(亚高温高光)下,培养5天后的相关指标数据如下表。
组别 | 温度/℃ | 光照强度/(μmol·m-2·s-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/ppm | Rubisco活性/(U·mL-1) |
CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降并不是由于气孔导度因素引起的,请说出理由
(4)D1蛋白是PSⅡ复合物的组成部分,对维持PSⅡ的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。某研究者利用番茄植株进行了三组实验,①组的处理为适宜温度和适宜光照处理番茄植株,②组的处理为

(2)以蓝细菌为研究模型,通过导入三种外源酶(ABC酶)基因,相关指标的检测结果见表和图2.
| 组别 | 导入基因 | NADPH含量(pmol) | ATP含量(μmol) | CO2固定速率(mg·g-1细胞干重·h-1) |
| 一 | 无 | 193.5 | 39.28 | 86 |
| 二 | A、B | 190.83 | 35.23 | 85 |
| 三 | A、B、C | 112.83 | 62.53 | 119 |

②结合表和图2分析,在蓝细菌中创建异丙醇合成途径能够提高光合速率的原因:
| 番茄大棚 | 草莓大棚 | |||||
| 空气质量 | 棚温(℃) | 光合有效辐射 | 光合速率 | 棚温(℃) | 光合有效辐射 | 光合速率 |
| 良 | 25.6 | 987 | 20.4 | 27.3 | 994 | 20.11 |
| 轻度污染 | 23.4 | 746 | 19.6 | 26.1 | 785 | 18.72 |
| 中度污染 | 23.5 | 477 | 17.1 | 26.1 | 428 | 17.41 |
| 重度污染 | 23.1 | 325 | 11.8 | 24.6 | 428 | 10.10 |
注:光合有效辐射指植物进行光合作用时,光合色素吸收并转化的太阳能。
(1)叶肉细胞吸收的CO2在
(2)据表分析,空气质量重度污染时,番茄大棚和草莓大棚应采取
(3)科研人员还探索持续光照是否导致植物有机物积累量增加。他们给予某种植物48小时持续光照,测定叶肉细胞中的淀粉积累量,结果如图1所示。据图分析,淀粉积累量的变化规律是

(4)为了解释(3)出现的实验结果,科研人员提出了两种假设。
假设一:当叶肉细胞内淀粉含量达到一定值后,淀粉的合成停止
假设二:当叶肉细胞内淀粉含量达到一定值后,淀粉的合成与降解同时存在。
为验证假设,科研人员测定了叶肉细胞的CO2吸收量和淀粉降解产物——麦芽糖的含量,结果如图2所示。实验结果支持假设


