组别 | 温度/℃ | 光照强度/(μmol·m-2·s-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/ppm | Rubisco活性/(U·mL-1) |
CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
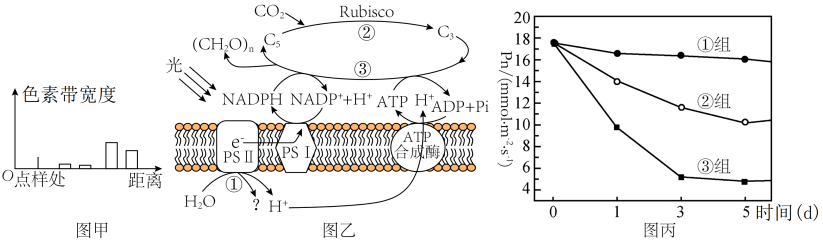
(1)分析图甲所示实验结果可知,含量最多的色素为
(2)PSII中的色素吸收光能后,将H2O分解为H+和
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降的原因
(4)D1蛋白是PSII复合物的组成部分,对维持PSII的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。已知药物SM可抑制D1蛋白的合成。为研究植物应对高温高光逆境时D1蛋白的变化机制,研究者利用番茄植株进行如下三组实验:①组在适宜温度、适宜光照下培养;③组在高温高光下培养并施加适量Sm(抑制D1蛋白合成的药物)。②组的处理方式是
材料一:某班学生进行新鲜番茄植株叶片色素的提取和分离实验,研磨时未加入CaCO3,实验结果如图甲所示。图乙是番茄植株进行光合作用的示意图,其中PSⅡ和PSⅠ是吸收、传递、转化光能的光系统。请回答下列问题:

(1)分析图甲所示实验结果可知,距离点样处距离最远的色素为
(2)PSⅡ中的色素吸收光能后,将H2O分解为H+和
材料二:某研究者测得番茄植株在CK条件(适宜温度和适宜光照)和HH条件(亚高温高光)下,培养5天后的相关指标数据如下表。
组别 | 温度/℃ | 光照强度/(μmol·m-2·s-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/ppm | Rubisco活性/(U·mL-1) |
CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
注:两组实验,除温度和光照有差异外,其余条件相同且适宜。
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降并不是由于
(4)D1蛋白是PSⅡ复合物的组成部分,对维持PSⅡ的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。某研究者利用番茄植株进行了三组实验,①组的处理同(2)中的CK,②组的处理为

表1 甲、乙两个水稻品种灌浆期和蜡熟期光合作用相关指标的比较
| 生长期 | 光补偿点(μmol·m-2·s-1) | 光饱和点(μmol·m-2·s-1) | 最大净光合速率(μmolCO2·m-2·s-1) | |||
| 甲 | 乙 | 甲 | 乙 | 甲 | 乙 | |
| 灌浆期 | 68 | 52 | 1853 | 1976 | 21.67 | 27.26 |
| 蜡熟期 | 75 | 72 | 1732 | 1365 | 19.17 | 12.63 |
(1)如果甲、乙两个水稻呼吸作用强度相同,则在灌浆期乙光补偿点低的直接原因是
(2)植物处于灌浆期,若此时光照强度为甲种水稻光补偿点,则在此光强下乙种水稻叶细胞中产生ATP的场所有
(3)从表中的数据推测,单位时间内,
(4)根据该实验的结果推测,从灌浆期到蜡熟期水稻最大净光合速率的变化可能与叶片的叶绿素含量变化有关。某研究小组设计实验验证该推测,请补充:
实验设计思路:
预期实验结果和结论:若
(2)马铃薯下侧叶片叶肉细胞中的叶绿体基粒可将光能转化为
(3)在一定浓度的CO2和30℃条件下,测定马铃薯和红薯在不同光照条件下的光合速率结果如下表。分析表中数据回答:
(注:细胞呼吸最适温度为30℃,光合作用最适温度为25℃)
| 光合速率与呼吸速率相等时光照强度(klx) | 光饱和时光照强度(klx) | 光饱和时CO2吸收量(mg/100cm²叶·小时) | 黑暗条件下CO2释放量(mg/100cm2叶·小时) | |
| 红薯 | 1 | 3 | 11 | 6 |
| 马铃薯 | 3 | 9 | 30 | 15 |
叶片 | 光补偿点(μmol.m2-.s-1) | 光饱和点(μmol.m-2.s-1) | 叶绿素a(mg.g-1) | 最大净光合速率 (nmolO2.g-1 min-1 ) |
新生阔叶 | 16.6 | 164.1 | 0.37 | 1017.3 |
繁殖期阔叶 | 15.1 | 266.0 | 0.73 | 1913.5 |
狭叶 | 25.6 | 344.0 | 0.54 | 1058. 2 |
(2)依据表中
(3)新生阔叶颜色呈浅黄色,欲确定其所含色素的种类,可用
(4)将新生阔叶由温度 18℃移至 26℃(为呼吸的最适温度)下,其光补偿点将
表外源NO对干旱胁迫下玉米幼苗光合作用和生长的影响
| 处理 | 气孔导度(mmolH2O·m-2·s-l) | 光合速率(μmolCO2m-2·s-l) | Fv/Fm | 根干重(g/plant) |
| 对照组 | 0.061 | 9.2 | 0.81 | 0.061 |
| PEG处理 | 0.010 | 3.1 | 0.68 | 0.063 |
| NO处理 | 0.089 | 9.1 | 0.81 | 0.068 |
| NO处理后再PEG处理 | 0.025 | 4.2 | 0.77 | 0.078 |
(备注:Fv/Fm代表叶绿体类囊体最大光能转化效率。)
回答下列问题:
(1)水既是光合作用
(2)为进一步研究PEG处理使玉米幼苗Fv/Fm下降的原因,利用试剂
(3)据表分析,使用外源NO处理可显著提高玉米幼苗在干旱胁迫时的气孔导度,直接促进卡尔文循环的
叶片 | 光补偿点(μmol•m ﹣2•s﹣1) | 光饱和点(μmol•m ﹣2•s﹣1) | 叶绿素 a(mg • g﹣1) | 最大净光合作用(nmol O2•g﹣1•min﹣1) |
新生阔叶 | 16.6 | 164.1 | 0.37 | 1017.3 |
繁殖期阔叶 | 15.1 | 266.0 | 0.73 | 1913.5 |
狭叶 | 25.6 | 344.0 | 0.54 | 1058.2 |
(1)据表分析,鼠尾藻从生长期进入繁殖期时,阔叶的光合作用强度
(2)依据表中
(3)新生阔叶颜色呈浅黄色,欲确定其所含色素的种类,可用
(4)将新生阔叶由温度 18℃移至 26℃(为呼吸的最适温度)下,其光补偿点将
| 不遮阴(对照) | 50%遮阴 | 75%遮阴 | 90%遮阴 | |
| 叶绿素a含量(mg·dm-2) | 2.03 | 3.42 | 4.15 | 4.04 |
| 叶绿素b含量(mg·dm-2) | 0.75 | 1.32 | 1.61 | 1.65 |
| 类胡萝卜素含量(mg·dm-2) | 1.05 | 1.49 | 1.71 | 1.72 |
| 最大光合速率(μmol·m-2·s-1) | 4.52 | 6.04 | 6.52 | 4.93 |
请分析回答下列问题
(1)据表推知,遮阴会导致香榧苗对
(2)在一定的范围内,香榧苗随着遮阴程度的增加光合速率有所提高,其原因可能是色素含量上升,利于对可见光中
叶绿素a mg/g | 叶绿素b mg/g | 光合速率 μmol/m2·s | 胞间CO2浓度 μmol/m2·s | 气孔导度 mol/m2·s | |
对照组 | 0.28 | 0.39 | 2.13 | 387.27 | 0.086 |
补水组 | 0.98 | 0.41 | 5.47 | 356.8 | 0.085 |
请回答:
(1)对照组的处理是
(2)叶绿素a具有吸收和转化光能的作用,叶绿素b具有吸收和传递光能的作用,表格数据说明生境陆化对秋茄叶绿素a的损害程度
(3)另有研究发现,给秋茄施用一定量的脱落酸类似物具有增强补水组实验效果的作用,推测其原因是由于脱落酸具有
| 组别 | 实验材料 | 实验试剂或处理方法 | 实验记录 |
| 1 | 水绵和好氧细菌 | 在没有空气的黑暗环境中,用极细的光束照射水绵 | 观察细菌分布的部位 |
| 2 | 叶片经饥饿处理后,一半遮光,一半曝光 | 碘蒸气处理 | 颜色反应 |
| 3 | 小球藻、14C标记的14CO2 | 提供适宜的光照和14CO2 | 追踪检测放射性14C的去向 |
| 4 | 菠菜叶 | 无水乙醇、层析液等 | 色素带的宽度、位置与颜色 |
| A.第1组实验说明了光合作用的场所是叶绿体,CO2的固定需要光照 |
| B.第2组实验可以观察到遮光部位变蓝,曝光部位不变蓝 |
| C.第3组实验在小球藻细胞中的三碳化合物、丙酮酸中也可能检测到14C |
| D.第4组实验中色素带的宽度反映的是色素的溶解度大小 |


