| TPTC浓度(ug/L) | 0 | 0.15 | 0.30 | 0.60 | 1.20 | 2.40 |
| 呼吸速率(10-12gC/cell·h) | 0.109 | 0.116 | 0.135 | 0.161 | 0.180 | 0.96 |
| 光合速率(10-12gC/cell·h) | 1.56 | 1.53 | 1.56 | 1.50 | 1.49 | 0.47 |
| 叶绿素a含量(10-12g/cell·h) | 0.136 | 0.142 | 0.163 | 0.182 | 0.203 | 0.073 |
(2)TPTC浓度过高,将
(3)据表中数据分析,在一定范围内,随TPTC浓度增大使等鞭金藻
(4)如坐标图纵坐标为细胞生长密度,横坐标为培养时间。图例浓度从左到右依次为:0mg/L,1.8mg/L,3.6mg/L,10.8mg/L,19.8mg/L。据图分析问题:
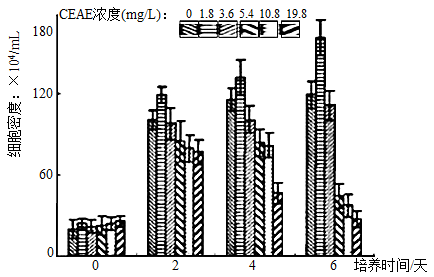
①在统计等鞭金藻细胞密度时,可用
②分析可知,CEAE对等鞭金藻生长的影响表现为“低促高抑”,浓度为
③每一个浓度均设置多次重复,遵循了实验设计的
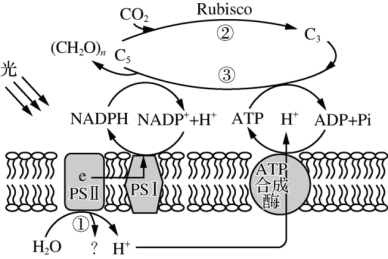
(1)PSⅡ中的色素吸收光能后,将H2O分解为H+和
(2)某研究者测得番茄植株在CK条件(适宜温度和适宜光照)和HH条件(亚高温高光)下,培养5天后的相关指标数据如下表。
| 组别 | 温度 /℃ | 光照强度/ (μmol·m-2·s-1) | 净光合速率/ (μmol·m-2·s-1) | 气孔导度/ (mmol·m-2·s-1) | 胞间CO2 浓度/ppm | Rubisco活性 /(U·mL-1) |
| CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
| HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
注:两组实验,除温度和光照有差异外,其余条件相同且适宜。
由表中数据可以推知,HH条件下番茄净光合速率的下降并不是由于
(3)D1蛋白是PSⅡ复合物的组成部分,对维持PSⅡ的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。某研究者利用番茄植株进行了三组实验,①组的处理同(2)中的CK,②组的处理为

(4)测定叶片中叶绿素含量时,采用“剪碎→研磨→过滤”的方法获取色素提取液,会导致色素提取不充分,其原因可能是
| 病级 | 病斑面积(%) | 叶绿素含量(%) | 净光合速率 [μmol. (m2.s)-1 | 气孔导度 [μmol. (m2s)-1] |
| 0 | 0.00 | 53.70 | 20.76 | 0.40 |
| 1 | 2.57 | 31.92 | 18.88 | 0.38 |
| 3 | 9.69 | 29.45 | 15.44 | 0.36 |
| 5 | 28.84 | 20.54 | 15.39 | 0.32 |
(1)提取黄瓜叶片中的光合色素时要使用
(2)在提取到黄瓜叶片光合色素后,可通过对比各组黄瓜叶片对某种颜色的光的吸收率来计算叶片中的叶绿素含量。为减少其他光合色素的干扰,“某种颜色的光”最可能是
(3)在实验过程中,黄瓜叶肉细胞产生O2的去向是
(4)某科研小组在晴朗的白天,分别对生长在强光下和弱光下的黄瓜幼苗吸收co,的速率(单位: gcm2-h-1)、气孔阻力(气孔开度减小,气孔阻力增大)等生理指标进行了测定,结果如下图所示。
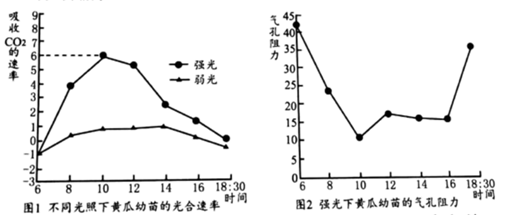
①若一天中温度不变,强光下黄瓜幼苗制造有机物的最大速率为
②若温度在一天中会发生变化,科研小组若要测定黄瓜幼苗一天中某一时段的光合速率,还要测定幼苗的
外源NaCl浓度( | 叶绿素a | 叶绿素b | 类胡萝卜素 |
50 | +28.98% | +3.49% | +7.58% |
100 | -11.05% | -17.04% | -18.95% |
200 | -25.81% | -41.47% | -30.61% |
400 | -48.01% | -62.21% | -58.89% |
 )
)注:“+”代表升高,“-”代表降低
请回答下列问题:
(1)实验室中分离绿叶中色素常用的方法是
(2)据表中数据推测,一定范围内的低浓度盐胁迫可以
(3)进一步研究发现,盐胁迫增大会造成叶绿体中的
(4)高盐环境还能影响植物细胞的渗透压,威胁植物生存。下列生理过程有利于植物抗盐的有
a.细胞膜上离子转运蛋白增加 b.从外界环境中吸收无机盐离子至液泡中
c.细胞质内合成可溶性有机小分子 d.细胞内产生一类具有调节渗透作用的蛋白质
采样日期 | 光合速率(相对值) | 叶绿素含量(mg/g) | ||
总量 | 游离型 | 结合型 | ||
7月1日 | 8.00 | 2.34 | 0.11 | 2.23 |
8月1日 | 7.80 | 2.30 | 0.12 | 2.18 |
9月1日 | 6.00 | 1.92 | 0.21 | 1.71 |
10月1日 | 4.20 | 1.58 | 0.29 | 1.29 |
11月1日 | 0.70 | 1.21 | 0.41 | 0.80 |
(1)为提取叶片中的叶绿素,研磨前在研钵中除加入剪碎的叶片外,还应加入
(2)结合表中数据,导致该叶片光合速率下降的主要原因是
(3)如图a、b是7月1日和11月1日该植物叶绿体亚显微结构照片,推测图

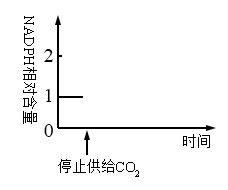

(1)水稻叶肉细胞进行光合作用时,能量转换过程是光能→
(2)PSII中的光合色素吸收光能后,一方面将水分解, 产生的电子经一系列传递体的传递,参与
| 对照组 | 实验组 | |
| Pn | 25.52±2.32 | 18.46±1.30 |
| Gs | 253.62±21.61 | 191.84±19.91 |
| 叶绿素含量(mg/g) | 35.12 | 26.35 |
①气孔导度的变化主要影响光合作用
②表中数据显示,空间搭载后DN416的
③研究人员同时研究了空间搭载对梗稻东农423(DN423) 光合作用的影响,发现其光合速率明显提升。这与DN416的研究结果相反,根本原因可能是空间搭载对水稻光合作用的影响与不同品种间
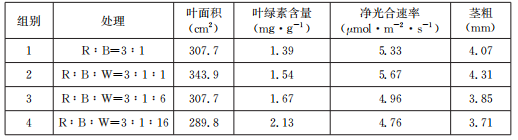
(1) 红(R)蓝(B)两种光能明显提高番茄幼苗的光合速率,原因是
(2) 该实验中,对照组为
(3) 第4组幼苗叶片中叶绿素含量最高,但净光合速率最低,原因可能是白光比例较高,导致叶绿体内一部分
(4) 据表中数据分析,可以得出的初步实验结论是
(5) 利用红蓝白(R∶B∶W=3∶1∶1)组合光处理后培育的番茄幼苗进行“绿叶中色素的提取和分离”实验,按规范操作提取出色素后,以无水乙醇作为层析液进行色素分离,发现滤纸条上各色素带的间距较小,最可能的原因是
表1 野生型和突变体的农艺性状
| 植株 | 株高/cm | 穗长/cm | 穗粒数/粒 | 结实率/% | 千粒重/g |
| YY | 109.4±4.2 | 21.9±0.5 | 169.9±14.5 | 75.8±3.2 | 28.4±0.5 |
| w08 | 104.9±3.7 | 21.0±0.6 | 159.1±16.6 | 37.8±1.7 | 23.2±0.5 |
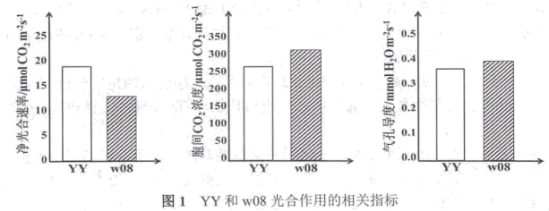
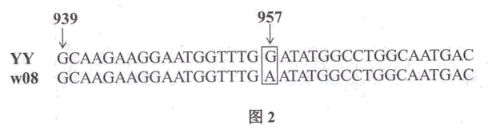
(1)据表1可知,w08的
(2)净光合速率除图中所示指标以外,还可以用
(3)研究发现w08中光合色素含量明显减少,导致光能转变成
(4)检测发现w08细胞内的叶绿素酸酯氧化酶基因(OsCAO1,与叶绿素合成相关)、脱镁叶绿酸a氧化酶基因(OsPAO,与叶绿素降解相关)及α/β水解酶家族蛋白基因(OsNYC3,与叶绿素降解相关)表达水平均显著下降。据此可推测:w08中叶绿素含量下降的主要原因是
光照 处理时间 | 净光合速率 Pn(μmol·m-2·s-1) | 蒸腾速率 Tr(mmol·m-2·s-1) | 气孔导度 Gs(mmol·m-2·s-1) | 胞间CO2浓度 Ci(μmol·mol-1) |
8 h·d-1 | 12.4 | 4.62 | 468.33 | 447.33 |
12 h·d-1 | 13.3 | 5.06 | 552.00 | 493.00 |
16 h·d-1 | 15.3 | 5.51 | 613.67 | 506.00 |
20 h·d-1 | 9.47 | 4.29 | 319.67 | 562.00 |
(1)常用
(2)请据表描述草莓净光合速率与光照处理时间的关系:
(3)蒸腾速率直接影响草莓叶片的温度,请结合表格数据,光照处理时间为20h·d-1时,净光合速率下降的可能原因是光照处理时间为20h·d-1时,气孔导度最低,
(1)研究人员首先研究了不同灌水量对该植物根、茎和叶形态变化的影响,结果如表1。
表1
茎粗/cm | 叶面积/m2 | 叶片数/片 | 新生气生根/条 | |
W70 | 54 | 172 | 6.88 | 5.44 |
W50 | 46 | 159 | 6.87 | 2.27 |
WCK | 47 | 163 | 6.76 | 9.32 |
实验结果表明,不同灌水量对
(2)研究人员接着研究了不同时期、不同灌水量下叶片净光合速率的变化情况,结果如下图,实验结果表明:
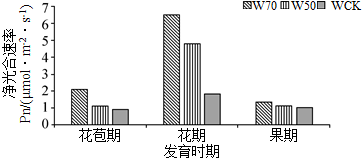
(3)研究人员进一步研究了不同灌水量对叶片胞间二氧化碳浓度和气孔导度的影响,发现W70条件下,叶片气孔导度变大,但胞间二氧化碳浓度不高,可能的原因是
(4)研究人员还研究了不同灌水量对不同发育时期的叶片光合色素含量的影响,提取并测定色素含量的具体操作如表2,具体结果如表3。
表2
实验操作 | 操作目的 |
每隔15天取完全展开的成熟叶片 | ① |
在叶片的上、中、下三个部位随机截取等量叶片并混合 | ② |
研磨叶片时加入无水乙醇 | ③ |
使用分光光度计进行测量样品吸光度 | ④ |
(注:将样品吸光度代入相关公式,可计算出样品中的物质含量)
表3
发育时期 | 处理 | 叶绿素a (mg·g-1) | 叶绿素b (mg·g-1) | 类胡萝卜素 (mg·g-1) |
花苞期 | W70 | 8.57 | 3.65 | 1.16 |
W50 | 8.19 | 3.57 | 0.9 | |
WCK | 5.49 | 1.37 | 0.7 | |
花 期 | W70 | 3.37 | 1.1 | 0.69 |
W50 | 2.85 | 0.84 | 0.39 | |
WCK | 2.46 | 0.77 | 0.36 | |
果期 | W70 | 14.12 | 4.06 | 2.17 |
W50 | 11.43 | 3.38 | 1.62 | |
WCK | 10.86 | 2.75 | 1.04 |
实验结果表明:在


