1 . 高中生物学实验中,下列实验操作能达到所述目标的是( )
| A.向泡菜坛盖边沿注满水可形成内部的无菌环境 |
| B.在目标个体集中分布的区域划定样方调查种群密度 |
| C.向血细胞计数板上滴加培养液后盖盖玻片可避免调查结果偏大 |
| D.用高浓度的蔗糖溶液处理洋葱鳞片叶内表皮细胞观察质壁分离 |
您最近半年使用:0次
2 . 高原鼢鼠是青藏高原优势地下啮齿动物(喜食杂类草植物根茎),每年3~6月进入繁殖期,其采食、挖掘洞道及推土造丘行为对高寒草地生态系统产生多种影响,为了更好的保护高寒草地生物多样性和科学防控草原鼠害,某科研小组对种群数量变化规律与环境因子的关联程度进行了探究。请回答下列问题。
(1)常采用土丘系数法(土丘系数=实捕鼢鼠数/土丘数)调查地下啮齿动物的种群数量。调查发现9个标准地(面积为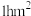 )的土丘数为a,鼢鼠种群密度为
)的土丘数为a,鼢鼠种群密度为______ 。调查期间,每天9:00~12:00在高原鼢鼠新推土丘位置附近利用探钎探明并剖开其活动洞道,并在洞道中布设10个活捕笼,该操作的意义是______ 。
(2)高原鼢鼠挖掘行为对高寒草地生态系统产生多种影响。一方面造成草地土壤更新有利于植物多样性的维持;另一方面,当高原鼢鼠种群密度超出______ 时导致草地生产力下降,生物多样性丧失,因其违背了______ 原理。
(3)由于高原鼢鼠种群特征对环境因子存在滞后效应,分别在春季(3~5月)、秋季(8~10月)进行研究,研究结果如下:表为2014~2019年间高原鼢鼠栖息地环境因子的变化,图为高原鼢鼠种群参数的变化
表高原鼢鼠栖息地环境因子的变化
①年际间种群密度、存活率整体呈______ 趋势,其原因可能是______ 。
②季节间存活率和种群补充量呈______ 趋势,其原因分别是______ 。
③春季杂类草生物量和高原鼢鼠种群特征有很高的关联度,分析原因______ 。
(1)常采用土丘系数法(土丘系数=实捕鼢鼠数/土丘数)调查地下啮齿动物的种群数量。调查发现9个标准地(面积为
)的土丘数为a,鼢鼠种群密度为(2)高原鼢鼠挖掘行为对高寒草地生态系统产生多种影响。一方面造成草地土壤更新有利于植物多样性的维持;另一方面,当高原鼢鼠种群密度超出
(3)由于高原鼢鼠种群特征对环境因子存在滞后效应,分别在春季(3~5月)、秋季(8~10月)进行研究,研究结果如下:表为2014~2019年间高原鼢鼠栖息地环境因子的变化,图为高原鼢鼠种群参数的变化
表高原鼢鼠栖息地环境因子的变化
2014 | 2015 | 2016 | 2017 | 2018 | 2019 | |
春季试验区平均地温 | 4.5 | 5 | 4 | 3.5 | 6 | 5 |
春季试验区雨量指数 | 18 | 10 | 29 | 24 | 6 | 50 |
秋季试验区平均地温 | 8 | 8.5 | 9 | 8 | 7.5 | 8.5 |
秋季试验区雨量指数 | 8.5 | 8 | 12 | 16 | 23 | 15 |
地上生物量 | 23.5 | 30 | 28 | 33 | 24 | 34 |
地下生物量 | 740 | 1000 | 600 | 540 | 860 | 780 |
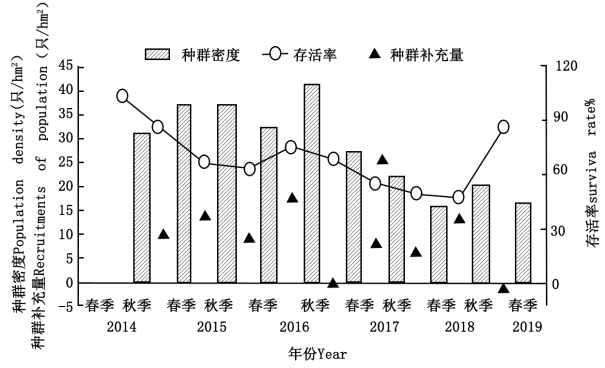
①年际间种群密度、存活率整体呈
②季节间存活率和种群补充量呈
③春季杂类草生物量和高原鼢鼠种群特征有很高的关联度,分析原因
您最近半年使用:0次
3 . 滨海某湿地森林植被退化,形成的裸滩被外来入侵植物互花米草占据。群落中芦苇、耐高盐的碱蓬和互花米草的相对多度(群落中某一种植物的个体数占该群落所有植物个体数的百分比)与入侵时间的关系如图1所示。研究发现,互花米草入侵后,导致图1两种植物相对多度发生变化的原因如图2所示。请回答下列问题:____ 。
(2)图1中乙植物是____ (填“芦苇”或“碱蓬”)。互花米草入侵的第1年至第5年甲种群密度变化的原因是____ 。
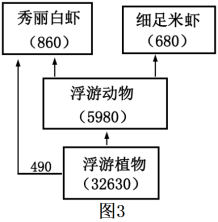
(3)该湿地某食物网如图3所示(图中数字为能量数值,单位为kJ/m2·a),该食物网中,第二营养级到第三营养级的能量传递效率为____ %(精确到小数点后一位)。已知这两种虾之间存在较为激烈的种间竞争,这种竞争会导致它们的生态位____ (填“重叠”或“分化”)。____ 。
(5)为防治互花米草的生态入侵,湿地修复过程中通常选择净化能力强的多种水生植物,使它们形成互利共存关系,这体现的生态学原理是____ 。
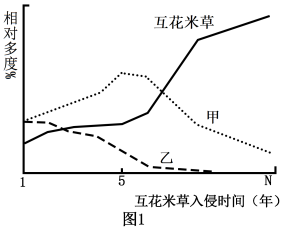

(2)图1中乙植物是
(3)该湿地某食物网如图3所示(图中数字为能量数值,单位为kJ/m2·a),该食物网中,第二营养级到第三营养级的能量传递效率为
(5)为防治互花米草的生态入侵,湿地修复过程中通常选择净化能力强的多种水生植物,使它们形成互利共存关系,这体现的生态学原理是
您最近半年使用:0次
4 . 榕属植物是我国南方最重要的园林景观植物。蓟马是榕属植物上的一种重要检疫性害虫,科研人员就该昆虫对榕属植物的选择行为进行了系列研究。请回答下列问题。
(1)蓟马的若虫和成虫群集于受害叶片上长成的虫瘿(植物组织因昆虫取食形成的瘤状物)里,吸食幼芽的汁液生存。对蓟马研究时,通常可用_____________ 法调查其种群密度。蓟马与榕属植物的种间关系是______________ ,蓟马同化的能量去向有______________ 。
(2)蓟马通过识别植物释放的一些挥发物来搜索适宜的榕属植物,该挥发物属于_____________ 信息。同时,榕属植物也可释放特定挥发物以驱避蓟马。这体现出信息传递在生态系统中的功能是______________ 。
(3)为研究蓟马对盆栽榕树挥发物的行为反应,研究人员使用图1所示Y形嗅觉仪开展实验。

①选取树龄、株高等条件相同且生长状态良好的三种盆栽榕树,用保鲜膜将育苗盆和泥炭土密封,分别放置于玻璃集气罐中,以收集三种榕树的_____________ 。
②按图1组装Y形嗅觉仪,各组件间分别通过软管连接。实验中将Y型管置于黑暗环境下,且正上方放置40W日光灯,以避免_____________ 。
③分别在两臂的样品瓶中放入待测气体,并将_____________ 处理12h的蓟马释放入Y形管基部管入口,统计爬向不同管的蓟马数量。每组处理观察60只蓟马,每只蓟马在实验中只使用1次。每测试10只更换Y型管并调换味源方位一次,目的是_____________ 。
④图2为实验结果,由此可知,蓟马对三种盆栽榕树挥发物的趋向性由大到小依次为_____________ 。

(4)上述系列研究,对种植榕属植物可借鉴性意义有_____________ 。
(1)蓟马的若虫和成虫群集于受害叶片上长成的虫瘿(植物组织因昆虫取食形成的瘤状物)里,吸食幼芽的汁液生存。对蓟马研究时,通常可用
(2)蓟马通过识别植物释放的一些挥发物来搜索适宜的榕属植物,该挥发物属于
(3)为研究蓟马对盆栽榕树挥发物的行为反应,研究人员使用图1所示Y形嗅觉仪开展实验。
①选取树龄、株高等条件相同且生长状态良好的三种盆栽榕树,用保鲜膜将育苗盆和泥炭土密封,分别放置于玻璃集气罐中,以收集三种榕树的
②按图1组装Y形嗅觉仪,各组件间分别通过软管连接。实验中将Y型管置于黑暗环境下,且正上方放置40W日光灯,以避免
③分别在两臂的样品瓶中放入待测气体,并将
④图2为实验结果,由此可知,蓟马对三种盆栽榕树挥发物的趋向性由大到小依次为
(4)上述系列研究,对种植榕属植物可借鉴性意义有
您最近半年使用:0次
2024-04-07更新
|
528次组卷
|
2卷引用:2024届江苏省苏锡常镇四市高三下学期教学情况调研(一) 生物试题
5 . 某野外调查小组对我国东部某群落进行深入调查,获得下面有关信息资料,请分析回答:
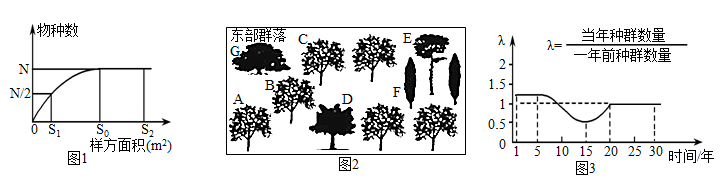
(1)图1是该小组调查获得的树林中物种数与面积的关系图,由图可知,调查该地区物种数的样方面积最好选___________ ,调查时,取样的关键是做到___________ 。
(2)该调查小组对该地区的树种形态特征进行了辨认和记录(如图2),并进行生物多样性研究:据图推测,树木A与树木B之间的差异属于___________ 多样性,树木A与树木D、E、G之间的差异属于___________ 多样性。
(3)图3是该调查小组从当地主管部门获得的某物种种群数量的变化图,据此分析该种群在第___________ 和第___________ 年之间是“J形”增长,第10-20年间,该种群的年龄结构是___________ ,在30年中种群数量最少开始于第___________ 年,第20~30间种群的增长率为___________ 。
(4)调查小组对群落中的几种植物的种群密度进行了连续五年的调查,调查的结果如下表所示(单位:株/m2)。
如果4种植物中有一种属于入侵物种,则该入侵物种最可能是___________ (填字母)。研究发现该群落中植物存在分层现象,该现象属于群落的___________ 结构,能显著提高群落_________ 。
(1)图1是该小组调查获得的树林中物种数与面积的关系图,由图可知,调查该地区物种数的样方面积最好选
(2)该调查小组对该地区的树种形态特征进行了辨认和记录(如图2),并进行生物多样性研究:据图推测,树木A与树木B之间的差异属于
(3)图3是该调查小组从当地主管部门获得的某物种种群数量的变化图,据此分析该种群在第
(4)调查小组对群落中的几种植物的种群密度进行了连续五年的调查,调查的结果如下表所示(单位:株/m2)。
年份 植物 | 2001 | 2002 | 2003 | 2004 | 2005 |
A | 10 | 9.2 | 8 | 6.8 | 5.5 |
B | 3 | 2.1 | 1.1 | 0.3 | 0.1 |
C | 0.3 | 0.6 | 1.1 | 1.5 | 2.3 |
D | 3 | 2 | 0.4 | 0 | 0 |
您最近半年使用:0次
6 . 某地区蝗虫在秋季产卵后死亡,以卵越冬。某年秋季降温提前,大量蝗虫在产卵前死亡,次年该地区蝗虫的种群密度明显下降。下列叙述错误的是( )
| A.调查蝗虫幼虫(跳蝻)的密度可以采用标志重捕法 |
| B.该地区蝗虫的卵、幼虫(跳蝻)、成虫构成一个种群 |
| C.蝗虫种群密度下降的原因是非密度制约因素导致出生率下降 |
| D.蝗虫的种群密度由于温度等随季节而变化,体现了群落的季节性 |
您最近半年使用:0次
7 . 下列关于种群、群落的叙述,错误 的是( )
| A.校园内蒲公英的随机分布,体现了群落的空间结构 |
| B.可利用粪堆数、活动痕迹对长臂猿进行野外种群数量监测 |
| C.干旱气候是影响蝗虫种群增长的非密度制约因素 |
| D.公园内不同鸟类生态位之间的差异提高了对环境资源的利用能力 |
您最近半年使用:0次
名校
8 . 照片重复取样法可推算雪豹种群密度。近日科研团队在祁连山某区域内布设了60台红外相机,连续拍摄了3个多月,确定研究区域约有20只雪豹。根据数学模型推算,估测在这个区域内每100km2生活有3只雪豹。下列叙述错误的是( )
| A.雪豹的种群密度调查还可采用粪便中微卫星DNA分子调查,不适合采用样方法 |
| B.决定雪豹种群密度的因素是出生率和死亡率、迁入率和迁出率 |
| C.若研究区域有120只雪豹,则估测该区域的种群密度是16只/100 km2 |
| D.利用照片重复取样法可用于调查一些大型稀少野生动物的种群密度 |
您最近半年使用:0次
9 . 东亚飞蝗具有异地迁飞、群集、生殖能力强大等特性,对农业生产具较大的危害性。为加强对东亚飞蝗的防治,避免蝗灾的发生,科学家对东亚飞蝗的种群特征、迁飞等进行了研究。请回答下列问题:
(1)蝗虫有散居型和群居型两种类型。散居和群居体现的是种群的___ 特征。散居型可向群居型转变,4-5只独居蝗虫聚集之后,可自发产生一种名为4-甲氧基苯乙烯的信息素,以吸引更多蝗虫聚集;随蝗虫密度的增加,释放的信息素含量也会迅速增加,进一步促进蝗虫的聚集,这体现出蝗虫的聚集过程属于___ 调节。群居时,蝗虫个体之间可通过后肢触碰传递信息,减少互相攻击、利于协调行动,这表明___ 离不开信息传递。
(2)鸭子是蝗虫的天敌,对散居状态的蝗虫控制效果较好。某科研小组通过在试验田放养鸭子研究对蝗虫的控制。表1为某段时间内植物和蝗虫的部分能量值(单位:104kJ),鸭子可捕食蝗虫和试验田中植物,该时间段内系统无有机物输出。
表1
鸭子的同化量为___ ,试验田中植物固定的能量中流入蝗虫与流入鸭子的能量比值为___ 。该研究表明,农业生产中采用牧鸭治蝗时还应考虑到___ 。
(3)为监测某地蝗虫的种群密度,可选择___ 法统计虫卵的数目。蝗虫喜欢在坚实的土地上产卵。若在蝗虫繁殖期人为疏松土壤,可影响蝗虫的___ ,而降低其种群密度。
(4)C3和C4植物体内的12C与13C的比值(δ)不同,测定东亚飞蝗翅中δ值,可反映其幼虫期的食物来源,进而判断其虫源地。研究者选择在某无可用耕地且不进行农事操作的试验岛屿上捕获东亚飞蝗,并检测成虫翅中δ值,进而推算出其幼虫期食物占比情况,结果如下图所示。试验岛屿相对位置和作物分布如表2所示。

表2
①试说明研究者选择该岛屿作为本实验观测点的原因是___ 。
②结合图、表信息,推测不同日期东亚飞蝗的迁飞方向为:___ 。
(1)蝗虫有散居型和群居型两种类型。散居和群居体现的是种群的
(2)鸭子是蝗虫的天敌,对散居状态的蝗虫控制效果较好。某科研小组通过在试验田放养鸭子研究对蝗虫的控制。表1为某段时间内植物和蝗虫的部分能量值(单位:104kJ),鸭子可捕食蝗虫和试验田中植物,该时间段内系统无有机物输出。
表1
| 项目 | 净同化量 | 呼吸消耗量 | 流向分解者 | 未利用 |
| 植物 | 108 | 75 | 21 | 58 |
| 蝗虫 | 7 | 10 | 1 | 3 |
(3)为监测某地蝗虫的种群密度,可选择
(4)C3和C4植物体内的12C与13C的比值(δ)不同,测定东亚飞蝗翅中δ值,可反映其幼虫期的食物来源,进而判断其虫源地。研究者选择在某无可用耕地且不进行农事操作的试验岛屿上捕获东亚飞蝗,并检测成虫翅中δ值,进而推算出其幼虫期食物占比情况,结果如下图所示。试验岛屿相对位置和作物分布如表2所示。
表2
| 区域 | 与试验岛屿的位置关系 | 主要作物 |
| A农业区 | 位于试验岛屿北侧约40km | 玉米、水稻 |
| B农业区 | 位于试验岛屿南侧约60km | 小麦、水稻 |
②结合图、表信息,推测不同日期东亚飞蝗的迁飞方向为:
您最近半年使用:0次
名校
10 . 生态学家Grime用三角形模型来解释植物的适应性(如图所示),该模型认为植物往往只有竞争能力强、抗生境严峻能力强、抗生境干扰能力强三种对策中的一种。Grime将生境的严峻定义为限制植物干重增长的外部强制因素,如光照、水分不足等,将生境干扰定义为破坏植物生物量的外力因素,如火烧、霜冻等。
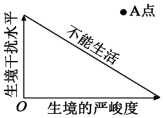
(1)Grime的三角形模型属于
(2)离乡间小路远近不同的地方,杂草的分布和种类不同,这体现了群落的
(3)我国库布齐沙漠日照强烈,地表温度高且降水蒸发快,结合Grime的三角形模型,在对该沙漠进行生态修复时,应选择种植抗生境严峻能力强的胡杨、沙柳等,这遵循了生态工程的协调和
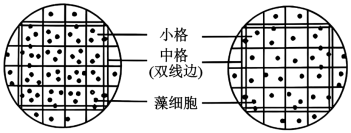
(4)研究者用血球计数板(1mm×1mm×0.1mm)对某单细胞藻进行计数了解治理效果。治理前后分别在相同样点取多个湖水样品混合后,稀释100倍进行计数,结果如图所示,左、右图分别为治理前、后某单细胞藻计数情况(一个计数室共25个中格)。治理后该藻种群密度减少了
(5)下表为太湖某区域生态系统的能量分析数据(单位:KJ·m-2·a-1),其中Ⅰ—Ⅳ表示食物链中不同的营养级,能量X为用于生长发育的能量。则表中生物类群V和能量Y分别表示的是
生物类群 | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ |
同化能量 | 871.27×105 | 141.1×105 | 15.91×105 | 0.88×105 | 211.85×105 |
能量X | 369.69×105 | 61.77×105 | 2.81×105 | 0.34×105 | 19.26×105 |
能量Y | 501.58×105 | 79.33×105 | 13.1×105 | 0.54×105 | 192.59×105 |
(6)从2020年10月起,政府对太湖实施十年禁渔。太湖禁渔期间
您最近半年使用:0次


