解题方法
1 . 已知果蝇眼色受一对等位基因控制。野生型果蝇眼色是暗红色,研究人员发现了一只新的亮红眼突变型雄果蝇,为探究亮红眼基因突变体的形成机制,设计了一系列实验(相关基因均不位于XY 染色体的同源区段)。
(1)亮红眼突变型雄果蝇与野生型雌果蝇杂交,F1果蝇自由交配,后代表现为_____ ,说明亮红眼是一种常染色体上的隐性突变,选择F2暗红眼果蝇自由交配,F1亮红眼果蝇的比例为_____ 。
(2)已知果蝇的长翅(V)对残翅(v)为显性,灰体(H)对黑檀体(h)为显性,棒眼(B)对正常眼(b)为显性,残翅(v)、黑檀体(h),正常眼(b)三个基因分别位于2号、3号和X染色体上,为探究亮红眼突变基因(用字母e表示)与上述三种基因的位置关系,以四种突变型果蝇(只有一对等位基因突变,其他性状均为野生型)为亲本进行杂交实验,方案及部分结果如下表所示。
①组合一果蝇杂交,F2中出现的结果为_____ 。
②组合三果蝇杂交,F2雌果蝇中b基因频率为_____ 。
(3)减数分裂时,雄果蝇3号染色体不发生互换,雌果蝇发生。为进一步确定亮红眼基因位于3 号染色体上,将组合二杂交产生的F1作_____ (填“母本”或“父本”)进行测交,若实验结果为_____ ,说明亮红眼基因位于3号染色体上。
(4)果蝇的眼色与色素合成细胞产生的眼黄素有关,眼黄素中色氨酸经过酶促反应合成。研究发现亮红眼果蝇眼睛中眼黄素显著偏低,而色氨酸酶促反应途径没有受到影响。由此推测,亮红眼基因与色氨酸_____ 有关。
(5)将野生型暗红眼和突变型亮红眼基因进行测序,下图为基因cDNA(转录的非模板链)的部分测序结果。_____ ,造成蛋白质翻译到第_____ 位氨基酸后提前终止(终止密码子: UAA、UAG、UGA)。
(1)亮红眼突变型雄果蝇与野生型雌果蝇杂交,F1果蝇自由交配,后代表现为
(2)已知果蝇的长翅(V)对残翅(v)为显性,灰体(H)对黑檀体(h)为显性,棒眼(B)对正常眼(b)为显性,残翅(v)、黑檀体(h),正常眼(b)三个基因分别位于2号、3号和X染色体上,为探究亮红眼突变基因(用字母e表示)与上述三种基因的位置关系,以四种突变型果蝇(只有一对等位基因突变,其他性状均为野生型)为亲本进行杂交实验,方案及部分结果如下表所示。
子代表现型 亲本组合 | 组合一: 亮红眼×残翅♀ | 组合二: 亮红眼×黑檀体♀ | 组合三: 亮红眼×正常眼♀ |
F1 | 暗红眼灰体 | ||
F2 | 暗红眼灰体:暗红眼黑檀体:亮红眼灰体=2:1:1 |
②组合三果蝇杂交,F2雌果蝇中b基因频率为
(3)减数分裂时,雄果蝇3号染色体不发生互换,雌果蝇发生。为进一步确定亮红眼基因位于3 号染色体上,将组合二杂交产生的F1作
(4)果蝇的眼色与色素合成细胞产生的眼黄素有关,眼黄素中色氨酸经过酶促反应合成。研究发现亮红眼果蝇眼睛中眼黄素显著偏低,而色氨酸酶促反应途径没有受到影响。由此推测,亮红眼基因与色氨酸
(5)将野生型暗红眼和突变型亮红眼基因进行测序,下图为基因cDNA(转录的非模板链)的部分测序结果。

您最近一年使用:0次
名校
2 . 番茄是世界主要蔬菜之一,为严格的自花授粉作物,杂种优势能极大提高番茄的产量、抗病及抗逆表现,因此番茄生产基本上都是应用杂交种。回答下列问题:
(1)科学家获得了位于4号染色体的ps-2基因隐性突变体,表现为雄性不育,在杂交育种时,选育雄性不育植株的优点是___________ 。
(2)番茄野生型为雄性可育,突变体甲和突变体乙均为雄性不育(均只有一对基因与野生型不同)。下表为3个不同番茄杂交组合及其子代的表型及比例。请回答:
根据杂交组合一和二可知,雄性可育性状是由___________ 性基因控制。根据杂交组合三,推测控制两个突变体的相关基因为___________ (填“等位基因”或“非等位基因”)。
(3)已知番茄的宽叶、窄叶由两对等位基因控制,现选择纯种宽叶番茄与窄叶番茄杂交,F1全部为宽叶,F1自交,F2中宽叶:窄叶为9:7
①F2中出现宽叶和窄叶比例为9:7的原因是___________ ,F2的窄叶有___________ 种基因型,若F2的宽叶自花传粉,则子代中窄叶的比例为___________ 。
②自然界中存在“自私基因”,即某一基因可以使同株的控制相对性状的另一基因的雄配子部分死亡,从而改变子代的表型比例。若宽叶、窄叶由一对等位基因(A、a)控制,F2中出现宽叶和窄叶的比例为9:7是“自私基因”作用的结果,则此比例出现的原因是:F1中携带___________ (填“A”或“a”)基因的雄配子,有___________ 的比例死亡。
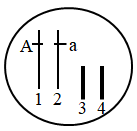
(4)我国科学家在番茄基因组中鉴定到154个在雄蕊中特异表达的基因,选取其中的一个基因S1STR1作为靶标基因(T表示)。利用CRISPR/Cas9基因编辑技术对番茄的S1STR1基因进行定向敲除获得雄性不育系(tt,绿色)。将正常功能的S1STR1基因(T)和控制花青素合成的SIANT1基因(A表示)连锁在一起,共同转回到雄性不育系中,从而获得了紫色的转基因保持系(如图)。关于该转基因保持系制备过程及在农业生产的优点,下列说法正确的是哪几项___________
(1)科学家获得了位于4号染色体的ps-2基因隐性突变体,表现为雄性不育,在杂交育种时,选育雄性不育植株的优点是
(2)番茄野生型为雄性可育,突变体甲和突变体乙均为雄性不育(均只有一对基因与野生型不同)。下表为3个不同番茄杂交组合及其子代的表型及比例。请回答:
| 组合序号 | 后代的表型及比例 | |
| 一 | 野生型×突变体甲 | 全为雄性可育(杂种1) |
| 二 | 野生型×突变体乙 | 全为雄性可育(杂种2) |
| 三 | 杂种1×杂种2 | 全为雄性可育 |
(3)已知番茄的宽叶、窄叶由两对等位基因控制,现选择纯种宽叶番茄与窄叶番茄杂交,F1全部为宽叶,F1自交,F2中宽叶:窄叶为9:7
①F2中出现宽叶和窄叶比例为9:7的原因是
②自然界中存在“自私基因”,即某一基因可以使同株的控制相对性状的另一基因的雄配子部分死亡,从而改变子代的表型比例。若宽叶、窄叶由一对等位基因(A、a)控制,F2中出现宽叶和窄叶的比例为9:7是“自私基因”作用的结果,则此比例出现的原因是:F1中携带
(4)我国科学家在番茄基因组中鉴定到154个在雄蕊中特异表达的基因,选取其中的一个基因S1STR1作为靶标基因(T表示)。利用CRISPR/Cas9基因编辑技术对番茄的S1STR1基因进行定向敲除获得雄性不育系(tt,绿色)。将正常功能的S1STR1基因(T)和控制花青素合成的SIANT1基因(A表示)连锁在一起,共同转回到雄性不育系中,从而获得了紫色的转基因保持系(如图)。关于该转基因保持系制备过程及在农业生产的优点,下列说法正确的是哪几项___________
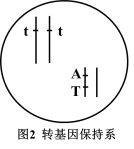
| A.转基因保持系通过杂交可产生雄性不育系又可产生转基因品系 |
| B.可通过幼苗颜色准确鉴定不育株用于杂交种子生产 |
| C.该技术用于杂交制种的不育系并不含任何转基因成分 |
| D.该研究策略易推广到其他蔬菜、花卉等园艺作物,具有广阔的应用前景 |
您最近一年使用:0次
2023-12-28更新
|
211次组卷
|
2卷引用:浙江省宁波市镇海中学2023-2024学年高三12月月考生物试题
3 . 野生型二倍体拟南芥基因型标记为WW,不含有潮霉素抗性基因H,表现为对潮霉素敏感。转入外源H基因获得纯合的转基因植株,一部分植株表现为对潮霉素抗性,基因型标记为RR;另一部分植株表现为对潮霉素敏感,基因型标记为SS。这些植株经染色体数目加倍处理后,获得四倍体拟南芥,基因型分别标记为WWWW、RRRR和SSSS。为研究转基因拟南芥对潮霉素敏感的原因,科研人员进行相关实验。
(1)选择上述植株作为亲本进行杂交实验,获得F1,F1自交得到F2,杂交结果如下表。
①由杂交一结果可知,_________ 为显性性状。
②杂交二中,F1植株的基因型标记为_________ 。杂交二的F1植株在减数分裂过程中,_________ 联会后平均分配,随机移向细胞两极,产生的三种生殖细胞基因型标记为_________ 。
③与杂交二的结果相比,杂交三的F2中敏感型植株所占比例高于预期。为解释上述现象,提出一种假设。
假设:四倍体中,潮霉素抗性植株RRRR的H基因处于激活状态能表达,潮霉素敏感植株SSSS中位于相同位点的H基因被甲基化而处于沉默状态不能表达。甲基化的H基因可能会诱导未被甲基化的H基因发生甲基化。
请依据上述假设,在答题卡上用遗传图解的方式,解释杂交三的F2中敏感型植株占1/4。
(2)研究者设法去除SS植株中H基因的甲基化,发现DNA甲基化抑制H基因转录,得出此结论的实验结果应为_________ 。
(3)请结合上述研究结果和表观遗传学相关知识,谈谈你对“基因与性状关系”的认识:_______ 。
(1)选择上述植株作为亲本进行杂交实验,获得F1,F1自交得到F2,杂交结果如下表。
杂交一 | 杂交二 | 杂交三 | ||||
P | RR | SS | RRRR | WWWW | RRRR | SSSS |
F1 | 均表现为潮霉素抗性 | |||||
抗性 | 敏感 | 抗性 | 敏感 | 抗性 | 敏感 | |
F2 | 3/4 | 1/4 | 35/36 | 1/36 | 3/4 | 1/4 |
②杂交二中,F1植株的基因型标记为
③与杂交二的结果相比,杂交三的F2中敏感型植株所占比例高于预期。为解释上述现象,提出一种假设。
假设:四倍体中,潮霉素抗性植株RRRR的H基因处于激活状态能表达,潮霉素敏感植株SSSS中位于相同位点的H基因被甲基化而处于沉默状态不能表达。甲基化的H基因可能会诱导未被甲基化的H基因发生甲基化。
请依据上述假设,在答题卡上用遗传图解的方式,解释杂交三的F2中敏感型植株占1/4。
(2)研究者设法去除SS植株中H基因的甲基化,发现DNA甲基化抑制H基因转录,得出此结论的实验结果应为
(3)请结合上述研究结果和表观遗传学相关知识,谈谈你对“基因与性状关系”的认识:
您最近一年使用:0次
2022-05-16更新
|
724次组卷
|
4卷引用:2023届浙江省高三选考模拟测试生物试题4
4 . 亚洲瓢虫的鞘翅呈现色彩丰富的斑点,鞘翅的黑缘型、均色型和黄底型分别由SASA、SESE和ss控制。为研究鞘翅色彩的遗传特点,用三组亚洲瓢虫进行杂交实验,F1自由交配得F2,其结果如下表:
(1)欲确定子一代新类型体细胞的染色体数目和形态特征,需对其进行_________ 。
(2)若仅考虑鞘翅的色彩斑点由SA、SE和s基因决定,则与之相关的瓢虫的基因型有_________ 种,表现型有_________ 种。
(3)根据甲、乙杂交组合的实验结果分析,子一代全为新类型,子二代出现图中不同表现形的现象称为_________ ,出现这种现象的原因是F1产生配子时,_________ 分离。
(4)丙组的子一代进行自由交配,在子二代中出现新类型的概率为_________ 。欲测定新类型三的基因型,可将其与表现型为_________ 瓢虫测交,若后代表现型及其比例为_________ ,则新类型三为杂合子。
(5)为了进一步明确鞘翅斑点的遗传特点,研究者又将黑缘型和均色型杂交,子代表现为两种亲本性状的嵌合体(如图所示),这种显性现象称为镶嵌显性。这种显性类型与人类AB血型的表现形式是有区别的,前者是将双亲的显性性状在_________ 表现,后者是将双亲的显性性状在子一代同一个体的相同细胞中表现。
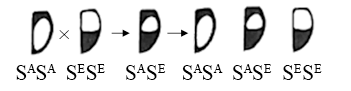
杂交组合 | 亲本 | 子一代 | 子二代 |
甲 | 均色型×黄底型 | 新类型一 | 均色型:新类型一:黄底型=1:2:1 |
乙 | 黑缘型×黄底型 | 新类型二 | 黑缘型:新类型二:黄底型=1:2:1 |
丙 | 新类型一×新类型二 | 黄底型:新类型一:新类型二:新类型三1:1:1:1 | ? |
(1)欲确定子一代新类型体细胞的染色体数目和形态特征,需对其进行
(2)若仅考虑鞘翅的色彩斑点由SA、SE和s基因决定,则与之相关的瓢虫的基因型有
(3)根据甲、乙杂交组合的实验结果分析,子一代全为新类型,子二代出现图中不同表现形的现象称为
(4)丙组的子一代进行自由交配,在子二代中出现新类型的概率为
(5)为了进一步明确鞘翅斑点的遗传特点,研究者又将黑缘型和均色型杂交,子代表现为两种亲本性状的嵌合体(如图所示),这种显性现象称为镶嵌显性。这种显性类型与人类AB血型的表现形式是有区别的,前者是将双亲的显性性状在
您最近一年使用:0次
2020-05-08更新
|
453次组卷
|
4卷引用:2020届浙江省台州市高三4月教学质量评估生物试题
2020届浙江省台州市高三4月教学质量评估生物试题(已下线)考点13 基因的分离定律-2020年【衔接教材·暑假作业】新高三一轮复习生物(人教版)(已下线)【浙江新东方】+2020年杭州市新东方学校高二上试卷【高中生物25】第一章 遗传的基本规律 第一节 孟德尔从一对相对性状的杂交实验中总结出分离定律
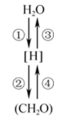
5 . 下图表示小麦叶肉细胞内两个重要的生理过程,表一表示小麦受镉浓度影响的相关测量数据,表二表示两种植物受光照强度影响的相关测量数据,请根据图表回答。
表一
(1)①过程的场所是__________ 。
(2)若该小麦长期缺Mg,则提取该小麦的色素经__________ 分离后,滤液细线自下而上第__________ 条色素带明显变窄。
(3)④过程产生的[H]来源是__________ (不考虑中间产物)。
(4)据表一分析,高剂量(≥1mg·L-1)镉会使气孔导度明显下降,而胞间二氧化碳浓度却增大,其主要原因是__________ 。
(5)表二取阳生和阴生两种长势相似的植物,分别放在两个相同的密闭透明玻璃瓶中,在适宜温度条件下,逐渐增加光照强度,测定放氧速率的数据如下表。请回答相关问题:
表二
①光强由200μmol光子/(100g·s)突然变为450μmol光子/(100g·s)时,阳生植物短期体内的三碳酸和RuBP变化分别为__________ 。
②该实验的目的是探究_________________________ 光合作用的影响。
③光饱和点时,阴生植物的呼吸熵为0.8(呼吸作用CO2的释放速率/呼吸作用O2的消耗速率),则该植物要从外界吸收CO2的速率是______ μmol/(100g·min)
镉浓度 (mg·L-1) | 气孔导度 (mmolCO2·m2·s-1) | 胞间CO2浓度 (μLm-2·s-1) | 净光合速率 (μmolCO2·m2·s-1) |
0 | 154.75 | 256.50 | 11.05 |
0.01 | 133.50 | 264.50 | 9.07 |
0.1 | 141.50 | 236.75 | 12.02 |
1 | 121.00 | 277.00 | 8.37 |
10 | 93.75 | 355.00 | 3.52 |
(1)①过程的场所是
(2)若该小麦长期缺Mg,则提取该小麦的色素经
(3)④过程产生的[H]来源是
(4)据表一分析,高剂量(≥1mg·L-1)镉会使气孔导度明显下降,而胞间二氧化碳浓度却增大,其主要原因是
(5)表二取阳生和阴生两种长势相似的植物,分别放在两个相同的密闭透明玻璃瓶中,在适宜温度条件下,逐渐增加光照强度,测定放氧速率的数据如下表。请回答相关问题:
光强μmol光 子/(100g·s) | 0 | 10 | 30 | 50 | 100 | 200 | 450 | 500 | |
放氧速率μ molO2/(100g·s) | 植物A | -20 | -10 | -5 | -1 | 5 | 15 | 29 | 28 |
植物B | -2 | -0.5 | 1.5 | 3 | 6 | 10 | 10 | 10 |
①光强由200μmol光子/(100g·s)突然变为450μmol光子/(100g·s)时,阳生植物短期体内的三碳酸和RuBP变化分别为
②该实验的目的是探究
③光饱和点时,阴生植物的呼吸熵为0.8(呼吸作用CO2的释放速率/呼吸作用O2的消耗速率),则该植物要从外界吸收CO2的速率是
您最近一年使用:0次
2020-09-25更新
|
216次组卷
|
2卷引用:2020届浙江省金华市东阳市高三6月模拟生物试题
解题方法
6 . 光抑制是指当植物吸收的光能超过其所需而导致光合速率下降的现象。光合复合体Ⅱ是光反应中吸收、传递并转化光能的一个重要场所,D1是其核心蛋白。在光能过剩时活性氧(ROS)的大量积累可直接破坏D1蛋白且抑制其合成,被破坏的D1降解后,空出相应的位置,新合成的D1占据相应位置,光系统Ⅱ得以修复。为探究高温胁迫对植物光合速率的影响机理,研究者进行了如下实验:
实验一:以野生型番茄植株为实验材料进行探究,在实验第3天时测定相关实验数据,如表所示(R酶参与碳反应)
(1)光合复合体Ⅱ位于叶绿体的_____ ,该复合体由蛋白质、电子传递体和_____ 组成,其吸收的能量用于光反应阶段。
(2)叶肉细胞吸收的CO₂,在_____ 被固定形成三碳酸,接受_____ 释放的能量,经过一系列的反应转化为糖类,并进一步合成淀粉。
(3)由表中数据可知,CO₂浓度_____ (是/不是)限制乙组番茄植株光合速率的重要因素,理由是_____ 。
(4)高温胁迫下番茄植株光反应速率减慢,分析其原因可能是:
①_____
②_____
实验二:已知叶绿素酶(CLH)能促进被破坏的D1降解。研究者以野生型番茄植株和CLH基因缺失的突变体植株为实验材料进行相关实验,测得实验结果如下图曲线所示(注:第1天之前为25℃,第1~3天为40℃,第4~5天为25℃)。_____ 。
(6)研究者补充设计以_____ 的突变型番茄植株为实验材料重复实验二,实验结果为上述结论提供支持证据。
实验一:以野生型番茄植株为实验材料进行探究,在实验第3天时测定相关实验数据,如表所示(R酶参与碳反应)
| 组别 | 温度 | 气孔导度 (mol·m²·s¹) | O₂释放速率(μmol·m²·s¹) | 胞间CO₂浓度(μmol·mol-¹) | R酶活性 (U·ml-¹) |
| 甲 | 25℃ | 99.2 | 11.8 | 282 | 172 |
| 乙 | 40℃ | 30.8 | 1.1 | 403 | 51 |
(1)光合复合体Ⅱ位于叶绿体的
(2)叶肉细胞吸收的CO₂,在
(3)由表中数据可知,CO₂浓度
(4)高温胁迫下番茄植株光反应速率减慢,分析其原因可能是:
①
②
实验二:已知叶绿素酶(CLH)能促进被破坏的D1降解。研究者以野生型番茄植株和CLH基因缺失的突变体植株为实验材料进行相关实验,测得实验结果如下图曲线所示(注:第1天之前为25℃,第1~3天为40℃,第4~5天为25℃)。
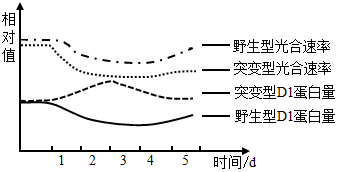
(6)研究者补充设计以
您最近一年使用:0次
2023-12-23更新
|
417次组卷
|
2卷引用:浙江省绍兴市嵊州市2023-2024学年高三上学期选考科目调研生物学科试题
7 . 某植株花色受两对基因A、a和B、b共同决定,基因A控制黄花,a控制蓝花,且当b基因纯合时遮盖基因A和a的表达,表现为红花。现用纯系植株进行杂交实验,得到如下实验结果。分析回答:
(1)基因A、a和B、b的遗传遵循_____ 定律,实验一中F1自交出现3/4黄花和1/4蓝花的原因是_____ 。
(2)实验二中红花亲本的基因型是_____ ,F2黄花个体中杂合子占_____ 。
(3)实验三中F2红花个体基因型有_____ 种,F2黄花个体间随机交配,后代表型及比例为_____ 。
(4)现有上述实验一中F1黄花植株经遗传改造得到的AaDH的黄花抗虫植株,已知A、a分别位于1号和2号染色体。D和H为外源导入的两个基因,已知这两个基因位于图示染色体上,且D基因位于3号染色体,同时具有D和H这两个基因的植株才表现为抗虫,现欲探究H基因的位置:
请完善实验思路,预测实验结果并分析讨论。
(注:每个植株的生殖力相同,且子代的存活率相同;D和H基因的插入不破坏其他基因:实验的具体操作不作要求)
实验思路:
第一步:选择_____ 植株自交获得子代:
第二步:观察记录子代表型及数量,并统计分析。
预测实验结果并分析讨论:
Ⅰ:若统计子代的表型及其比例为黄花抗虫∶黄花不抗虫∶蓝花不抗虫=9∶3∶4,则H基因位于_____ 染色体;
Ⅱ:若统计子代的表型及其比例为_____ ,则H基因位于2号染色体;
Ⅲ:若统计子代的表型及其比例为_____ ,则H基因位于4号染色体;
Ⅳ:若统计子代的表型及其比例为黄花抗虫∶黄花不抗虫∶蓝花抗虫∶蓝花不抗虫=9∶3∶3∶1,则H基因位于_____ 染色体。
| 实验组别 | 亲本 | F1表型 | F2表型 |
| 一 | 黄花×蓝花 | 黄花 | 3/4黄花1/4蓝花 |
| 二 | 黄花×红花 | 黄花 | 3/4黄花1/4红花 |
| 三 | 红花×蓝花 | 黄花 | 9/16黄花3/16蓝花1/4红花 |
(2)实验二中红花亲本的基因型是
(3)实验三中F2红花个体基因型有
(4)现有上述实验一中F1黄花植株经遗传改造得到的AaDH的黄花抗虫植株,已知A、a分别位于1号和2号染色体。D和H为外源导入的两个基因,已知这两个基因位于图示染色体上,且D基因位于3号染色体,同时具有D和H这两个基因的植株才表现为抗虫,现欲探究H基因的位置:
请完善实验思路,预测实验结果并分析讨论。
(注:每个植株的生殖力相同,且子代的存活率相同;D和H基因的插入不破坏其他基因:实验的具体操作不作要求)
实验思路:
第一步:选择
第二步:观察记录子代表型及数量,并统计分析。
预测实验结果并分析讨论:
Ⅰ:若统计子代的表型及其比例为黄花抗虫∶黄花不抗虫∶蓝花不抗虫=9∶3∶4,则H基因位于
Ⅱ:若统计子代的表型及其比例为
Ⅲ:若统计子代的表型及其比例为
Ⅳ:若统计子代的表型及其比例为黄花抗虫∶黄花不抗虫∶蓝花抗虫∶蓝花不抗虫=9∶3∶3∶1,则H基因位于
您最近一年使用:0次
2023-04-27更新
|
376次组卷
|
2卷引用:浙江省稽阳联谊学校2022-2023学年高三下学期4月联考生物试题
8 . 为了研究小鼠肥胖形成的内在机制,以小鼠为实验材料,进行了一系列的观察和探究实验。
(1)通过观察发现小鼠脂肪组织里穿插着一些交感神经,这些交感神经末梢会通过_____ 的方式释放神经递质—去甲肾上腺素。该递质能与脂肪细胞表面的____ 结合,进而促进细胞内的脂肪分解,释放能量。
(2)在显微镜下发现交感神经周边存在特殊的巨噬细胞,称为交感神经相关巨噬细胞(SAM),SAM具有许多伪足,能包裹住未鞘化的交感神经元轴突。为研究高脂食物投喂造成的肥胖对小鼠SAM细胞数量的影响,对两组肥胖小鼠进行投喂。实验组投喂____ 食物,对照组投喂____ 食物。在其他条件适宜的环境中培养一段时间,统计小鼠____ 。结果表明高脂食物投喂会促进肥胖小鼠SAM细胞的增殖。
(3)研究发现SAM细胞的细胞膜上具有去甲肾上腺素转运载体,为进一步明确SAM细胞与小鼠肥胖的关系,对普通小鼠和肥胖小鼠进行如下实验,结果如下表所示
不同处理条件下小鼠细胞脂肪分解速度
表中数据显示,注射去甲肾上腺素载体抑制剂能_______ 脂肪细胞脂肪分解速度。综上所述,小鼠肥胖形成的机制是__________________ 。
(1)通过观察发现小鼠脂肪组织里穿插着一些交感神经,这些交感神经末梢会通过
(2)在显微镜下发现交感神经周边存在特殊的巨噬细胞,称为交感神经相关巨噬细胞(SAM),SAM具有许多伪足,能包裹住未鞘化的交感神经元轴突。为研究高脂食物投喂造成的肥胖对小鼠SAM细胞数量的影响,对两组肥胖小鼠进行投喂。实验组投喂
(3)研究发现SAM细胞的细胞膜上具有去甲肾上腺素转运载体,为进一步明确SAM细胞与小鼠肥胖的关系,对普通小鼠和肥胖小鼠进行如下实验,结果如下表所示
不同处理条件下小鼠细胞脂肪分解速度
组别 | 实验材料 | 实验操作 | 脂肪细胞脂 肪分解速度 相对值 |
| 实验一 | 肥胖小鼠 | 注射生理盐水 | 70 |
| 实验二 | 肥胖小鼠 | 注射生理盐水+去甲肾上腺素载体抑制剂 | 90 |
| 实验三 | 普通小鼠 | 注射生理盐水 | 100 |
| 实验四 | 普通小鼠 | 注射生理盐水+去甲肾上腺素载体抑制剂 | 110 |
您最近一年使用:0次
2021-11-06更新
|
105次组卷
|
2卷引用:浙江省诸暨市2018-2019学年高三上学期期末生物试题
名校
解题方法
9 . 材料一:某班学生进行新鲜番茄植株叶片色素的提取和分离实验,研磨时未加入CaCO3,实验结果如图甲所示。图乙是番茄植株进行光合作用的示意图,其中PSII和PSI是吸收、传递、转化光能的光系统。
注:两组实验,除温度和光照有差异外,其余条件相同且适宜。
(1)分析图甲所示实验结果可知,含量最多的色素为____ ,可见光通过三棱镜后,照射到材料一中的色素提取液,发现其与正确操作下获得的色素提取液的吸收光谱差异最大在于____ 光。
(2)PSII中的色素吸收光能后,将H2O分解为H+和____ 。图乙中____ 为过程③供能,其中H+在____ 积累,从而推动ATP的合成。
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降的原因____ 。此条件下的短时间内光反应产物NADPH和ATP在叶绿体中的含量____ (选填“增加”、“减少”或“不变”)。
(4)D1蛋白是PSII复合物的组成部分,对维持PSII的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。已知药物SM可抑制D1蛋白的合成。为研究植物应对高温高光逆境时D1蛋白的变化机制,研究者利用番茄植株进行如下三组实验:①组在适宜温度、适宜光照下培养;③组在高温高光下培养并施加适量Sm(抑制D1蛋白合成的药物)。②组的处理方式是____ 。其他条件相同且适宜,连续5天每天定期测定各组番茄植株的净光合速率(Pn),结果如丙图,预测结果为三组D1蛋白含量从高到低依次是:____ 。
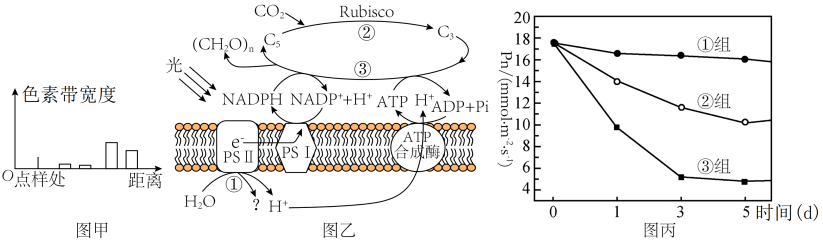
组别 | 温度/℃ | 光照强度/(μmol·m-2·s-1) | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/ppm | Rubisco活性/(U·mL-1) |
CK | 25 | 500 | 12.1 | 114.2 | 308 | 189 |
HH | 35 | 1000 | 1.8 | 31.2 | 448 | 61 |
(1)分析图甲所示实验结果可知,含量最多的色素为
(2)PSII中的色素吸收光能后,将H2O分解为H+和
(3)由表中数据可以推知,HH条件下番茄净光合速率的下降的原因
(4)D1蛋白是PSII复合物的组成部分,对维持PSII的结构和功能起重要作用,且过剩的光能可使D1蛋白失活。已知药物SM可抑制D1蛋白的合成。为研究植物应对高温高光逆境时D1蛋白的变化机制,研究者利用番茄植株进行如下三组实验:①组在适宜温度、适宜光照下培养;③组在高温高光下培养并施加适量Sm(抑制D1蛋白合成的药物)。②组的处理方式是
您最近一年使用:0次
2024-01-13更新
|
405次组卷
|
4卷引用:2024届浙江省宁波市奉化中学高考考前最后一卷生物试题
10 . 生物工程相关题目如下:
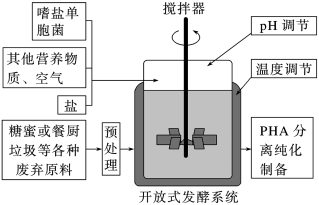
(一)中国科学家运用合成生物学方法构建了一株嗜盐单胞菌H以糖蜜(甘蔗榨糖后的废弃液,含较多蔗糖)为原料,在实验室发酵生产PHA等新型可降解材料,期望提高甘蔗的整体利用价值。工艺流程如图所示。_____________ ,并不断提高其浓度,经多次传代培养以获得目标菌株。培养过程中定期取样并用________ 方法进行菌落计数评估菌株增殖状况。此外,选育优良菌株的方法还有_______________ 等。(答出两种方法即可)
(2)基于菌株H嗜盐、酸碱耐受能力强等特性,研究人员设计了一种不需要灭菌的发酵系统,其培养基浓度设为60g/L,pH为10,菌株H可正常持续发酵60d以上。该系统不需要灭菌的原因是_______________ 。(答出两点即可)
(3)研究人员在工厂进行扩大培养,在适宜的营养物浓度、温度、pH条件下发酵,结果发现发酵液中菌株H细胞增殖和PHA产量均未达到预期,并产生了少量乙醇等物质,说明发酵条件中_______________ 可能是高密度培养的限制因素。
(4)菌株H还能通过分解餐厨垃圾(主要含蛋白质、淀粉、油脂等)来生产PHA,说明其能分泌_______________ 。
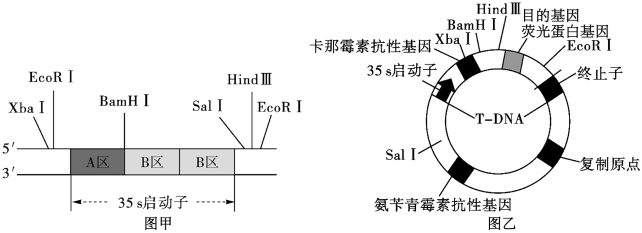
(二)35s启动子是广泛应用于转基因植物中的重要组件之一。一个35s启动子包括1个A区和2个B区。图甲表示35s启动子及其附近区域分布的限制酶识别位点,已知不同限制酶切割后得到的黏性末端各不相同。研究人员拟构建含有两个35s启动子串联的转基因苜蓿。图乙表示待转入第二个35s启动子的重组质粒的构成组件及其上的限制酶识别位点。
(注:图乙中的卡那霉素抗性基因可以在原核生物中表达)_______________ 。
(6)利用PCR技术扩增35s启动子过程中,每次循环中温度最低的步骤是_______________ 。根据限制酶识别位点,为了让第二个35s启动子准确串联在待转入的重组质粒上,应选择_______________ 限制酶切割甲图DNA片段。
(7)为了筛选出正确串联了两个35s启动子的新重组质粒菌落,研究人员用含有不同抗生素的平板进行筛选,得到①②③三类菌落,其生长状况如下表(+代表生长,-表示不生长)。根据表中结果判断,应选择的菌落是_______________ (填①、②或③)类,另外两类菌落质粒导入情况分是_______________ 、_______________ 。若新获得的转基因苜蓿中目的基因的表达量大幅增加,可以说明串联两个35s启动子能_______________ 。
(一)中国科学家运用合成生物学方法构建了一株嗜盐单胞菌H以糖蜜(甘蔗榨糖后的废弃液,含较多蔗糖)为原料,在实验室发酵生产PHA等新型可降解材料,期望提高甘蔗的整体利用价值。工艺流程如图所示。
(2)基于菌株H嗜盐、酸碱耐受能力强等特性,研究人员设计了一种不需要灭菌的发酵系统,其培养基浓度设为60g/L,pH为10,菌株H可正常持续发酵60d以上。该系统不需要灭菌的原因是
(3)研究人员在工厂进行扩大培养,在适宜的营养物浓度、温度、pH条件下发酵,结果发现发酵液中菌株H细胞增殖和PHA产量均未达到预期,并产生了少量乙醇等物质,说明发酵条件中
(4)菌株H还能通过分解餐厨垃圾(主要含蛋白质、淀粉、油脂等)来生产PHA,说明其能分泌
(二)35s启动子是广泛应用于转基因植物中的重要组件之一。一个35s启动子包括1个A区和2个B区。图甲表示35s启动子及其附近区域分布的限制酶识别位点,已知不同限制酶切割后得到的黏性末端各不相同。研究人员拟构建含有两个35s启动子串联的转基因苜蓿。图乙表示待转入第二个35s启动子的重组质粒的构成组件及其上的限制酶识别位点。
(注:图乙中的卡那霉素抗性基因可以在原核生物中表达)
(6)利用PCR技术扩增35s启动子过程中,每次循环中温度最低的步骤是
(7)为了筛选出正确串联了两个35s启动子的新重组质粒菌落,研究人员用含有不同抗生素的平板进行筛选,得到①②③三类菌落,其生长状况如下表(+代表生长,-表示不生长)。根据表中结果判断,应选择的菌落是
平板类型 | ① | ② | ③ |
无抗生素 | + | + | + |
卡那霉素 | · | , | + |
氨苄青霉素 | . | + | + |
卡那霉素+氨苄青霉素 | - | - | - |
您最近一年使用:0次
2023-12-25更新
|
378次组卷
|
2卷引用:2024届浙江省三校高三上学期联考生物试题


