净光 合速率(μmol·m-2·s-1) | 气孔 导度(mol·m-2·s-1) | 胞间CO2 浓度(μmol·mol-1) | CAT活性 (U·g-1FW·min-1) | 光反应转化的光 能/吸收的光能 | |
常温组 | 24.3 | 570 | 152 | 65 | 0.84 |
高温组 | 12.7 | 403 | 259 | 47 | 0.72 |
(1)某同学认为高温组净光合速率降低的原因是气孔导度降低,导致光合作用所需CO2减少,进而限制了暗反应,降低了光合速率。据表分析他的观点
(2)研究者对两组玉米叶片的叶绿素相对含量进行检测,结果如图。
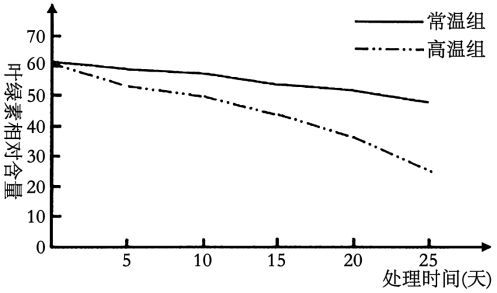
(3)综合以上结果,解释高温条件下叶片光合速率较低的原因可能是
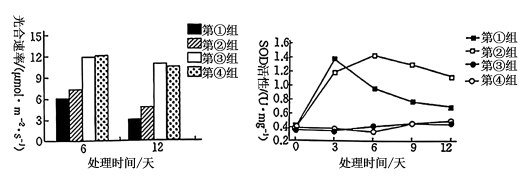
(2)低温胁迫诱发光抑制的植物的CO2固定速率降低,原因可能是
(3)结合实验结果分析,植物生长调节剂甲可
组别 | 处理 | 株高/cm | 鲜重/mg | 干重/mg | 根系活力/(μg·g-1·h-1) |
CK1 | 正常 | 26.51 | 297.71 | 62.28 | 98.25 |
CK2 | 干旱 | 19.50 | 231.72 | 49.37 | 54.60 |
T1 | 干旱+0.01mg/L BR | 24.77 | 269.13 | 58.86 | 97.07 |
T2 | 干旱+0.1mg/L BR | 29.93 | 316.63 | 62.18 | 104.61 |
T3 | 干旱+1mg/L BR | 25.20 | 288.78 | 57.50 | 97.73 |
(2)植物光合作用与植物的生长密切相关,为了进一步探究BR对植物光合作用的影响,科研人员进行了如下实验。
组别 | 处理 | 叶绿素a/(mg/L) | 叶绿素b/(mg/L) | 类胡萝卜素/(mg/L) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/(μmol/mol) | 净光合作用速率/(μmol·m-2·s-1) |
CK1 | 正常 | 1.68 | 0.71 | 0.34 | 0.048 | 322.31 | 1.48 |
CK2 | 干旱 | 1.32 | 0.38 | 0.46 | 0.021 | 346.58 | 0.85 |
T2 | 干旱+BR | 1.61 | 0.62 | 0.29 | 0.031 | 316.68 | 1.45 |
(3)为了进一步探究叶面营养剂及其与BR配合对羊草干旱胁迫的影响,还需要增加
(1)植物绿叶中的叶绿素通常可用
(2)生长素和油菜素内酯协同参与了植物的遮阴反应。研究人员探究了phyA突变体拟南芥(缺乏phyA)通过影响哪种激素信号的传导过程来抑制遮阴反应,相关实验内容如下:
组别 | 实验操作 | 实验现象和结果 | 实验结论 |
一 | 对野生型拟南芥和phyA突变体 拟南芥施用等量适量的NPA,在 遮阴条件下培养一段时间后,下 胚轴伸长量分别记为Al和A2 | 伸长量的大小关系 为:A1① 野生型的遮阴反应被NPA抑 制,而phyA突变体相对不敏感 | ④ |
二 | ② 一段时间后,下胚轴伸长量分 别记为B1和B2 | 伸长量的大小关系为:B1大 于B2,说明③ |
(3)为保证作物产量,请根据上述信息,提出一项耐阴、耐密植作物的培育建议:
(1)叶绿素在光合作用中起到
(2)叶绿体是光合作用的场所,某实验小组将叶绿体和相关化学物质在抽去空气的溶液中进行了如下研究:
组别 | 叶绿体类型 | 加入的物质 | 条件 | 溶液中的物质及颜色 |
甲组 | 完整叶绿体 | NADP+溶液 | 适宜温度和光照等条件 | 微量O2 |
乙组 | 叶绿体破裂后的匀浆 | NADP+溶液 | 微量O2和NADPH | |
丙组 | 完整叶绿体 | DCPIP溶液 | 微量O2,溶液为蓝色 | |
丁组 | 叶绿体破裂后的匀浆 | DCPIP溶液 | ? |
①上表中丁组溶液中的物质及颜色是
②下列关于本实验的分析错误的是哪几项
A.甲组和乙组比较,说明NADP+不能穿过叶绿体的双层膜
B.该实验说明产生的
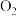 中的氧元素一定来源于
中的氧元素一定来源于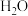 ,而不来源于
,而不来源于
C.将实验后的甲组在黑暗下处理一段时间,会有
生成D.离体的叶绿体基质中添加ATP、NADPH和
后,可完成碳反应Ⅱ.光是植物整个生命周期中许多生长发育过程的重要调节信号。我国科研人员对蓝光和赤霉素在调节植物发育中的相互关系进行研究。
(3)在拟南芥中,赤霉素与细胞内的赤霉素受体结合形成复合物,该复合物与R蛋白结合使R蛋白降解,从而抑制相关基因的表达,引起细胞伸长、植株增高。用赤霉素处理野生型和蓝光受体缺失突变体拟南芥后,用蓝光照射,分别检测R蛋白的含量,结果如图1。据图1结果推测:“野生型+黑暗”组比“野生型+蓝光”植株
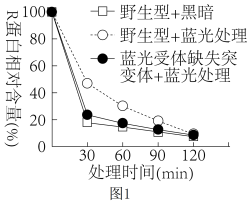
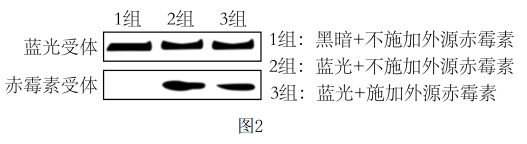
综合上述研究表明植物生长发育的调控是由基因表达调控、

(2)40天后突然解除遮阴,叶绿体中NADPH/NADP+、ATP/ADP的值会
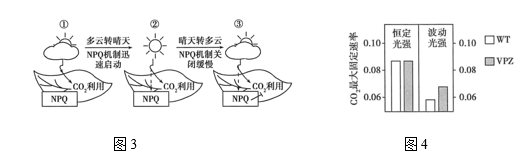
(3)长期生长在弱光下的植物叶片具有阴生叶的特点,遮阴解除当天突然暴露于强光下后,叶片发生的光抑制或光破坏严重。植物在长期进化过程中,其光合器官形成了一系列抵御强光破坏的光保护机制,如叶片和叶绿体运动相关的避光机制、非光化学淬灭机制(NPQ)、PSII损伤修复、活性氧自由基清除系统、光呼吸途径等。在NPQ的作用下多余的光能会以热能的形式散失。该机制的启动和关闭特点如图3所示,其中符号“ ┥”代表抑制作用。
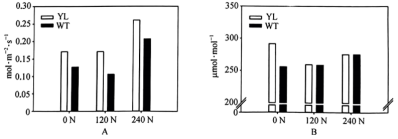
(4)科学研究者研发了一种转基因烟草(VPZ),相比野生烟草(WT)其在强光转为弱光后NPQ机制关闭的时间缩短。图4为分别在恒定光强和波动光强下测得的两种烟草的CO2最大固定速率。农作物一半的光合作用是在有各种阴影时进行的。根据上述信息推断,同样环境条件下VPZ的产量比WT的产量
(2)在饱和光强下,限制WT光合作用的主要内外因素分别是
(3)图示结果表明,在中氮与高氮处理下,YL相比WT的气孔导度大,但二者胞间CO2浓度却无显著差异,由此推断在中氮与高氮处理下,YL光合速率
(4)叶片中Rubisco含量高有利于提高光合速率,但合成Rubisco酶需要消耗大量的氮素。已知YL的Rubisco含量显著高于WT,这表明
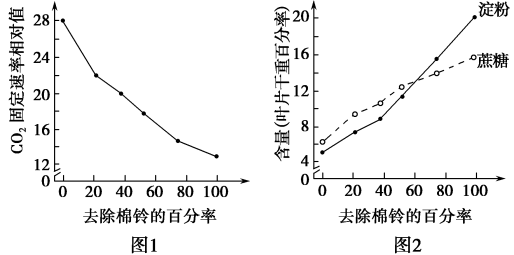
(1)绿叶通过气孔从外界吸收的CO2,在特定酶的作用下,在
(2)已知叶片光合产物会被运到棉铃等器官并被利用,据此分析图2中去除棉铃后,叶片中淀粉和蔗糖含量上升的原因是
(3)根据实验结果推测,叶片中光合产物的积累会
(4)为验证(3)的推测,可进行以下实验:去除植株上的棉铃并对部分叶片进行
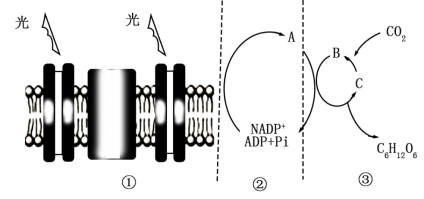
(2)当①中的光照突然减弱时,短时间内③中B、C的含量变化分别是
(3)③中催化CO2固定的酶(Rubisco)是一个双功能酶,CO2浓度高时,倾向于催化五碳化合物和CO2反应;O2浓度高时,倾向于催化五碳化合物和O2反应生成CO2(称为光呼吸)。因而当光呼吸较强时,叶肉细胞的光合速率会降低,据此分析原因是
处理 | 栽培密度 | 施氮量(kg·N·ha-1) | 叶绿素含量(mg·g-1·FW) | 产量(t·ha-1) | 光合速率(μmolCO2m-2s-1) | 氮肥利用率(kg·kg-1N) |
A | 100% | 0 | 3.29 | 7.29 | 20.89 | — |
B | 100% | 300 | 4.01 | 9.75 | 24.03 | 8.20 |
C | 120% | 270 | 3.58 | 10.75 | 23.88 | 12.81 |
(1)比较常规栽培与C组水稻的叶绿素含量差异时,提取色素过程中可添加
(2)与常规栽培B组相比,C组的水稻叶绿素含量下降,光反应中转移到
(3)有人认为上述表格结果也可以支持这样一个观点,即适当降低水稻叶绿素含量,可以减少农业生产对水体的富营养化污染。请阐述你的观点和理由


