解题方法
1 . 某种鸟的羽毛有覆羽和单羽是一对相对性状(基因为 A、a);B、b基因位于常染色体上,bb可使部分应表现为单羽的个体表现为覆羽,其余基因型无此影响。从群体中随机选取三对组合进行杂交实验,如下表所示。下列说法错误的是( )
| 杂交组合 | 子代 |
| 组合一:单羽雄鸟×覆羽雌鸟 | 覆羽雄鸟:单羽雌鸟=1:1 |
| 组合二:覆羽雄鸟×单羽雌鸟 | 全为覆羽 |
| 组合三:覆羽雄鸟×覆羽雌鸟 | 出现单羽雄鸟 |
| A.控制羽毛性状的基因位于Z染色体上,覆羽由Z染色体上的A基因控制 |
| B.组合一亲本基因型可能是bbZaZa、BBZAW |
| C.组合二子代覆羽雄鸟基因型是ZAZa或ZAZA |
| D.组合三子代单羽雄鸟基因型可能为bbZaZa |
您最近一年使用:0次
2 . 溶液培养法是用含有全部或部分营养元素的溶液栽培植物的方法。表一是小麦植株中占干重比例在0.01%以上的元素;表二是小麦培养液配方中部分成分(表中化合物在农业生产上应用称为化肥);表三是培养液中元素的质量浓度。请分析回答:
表一
表二
表三
(1)镁元素是构成____________ 必需元素,也是许多酶的活化剂。
(2)与表一相比,表三中缺少的元素有________ ,这些元素主要来自于小麦周围环境中的________________ 。
(3)小麦种子中蛋白质含量占11%左右,小麦面粉做成熟食后更容易消化吸收。从蛋白质结构分析,高温导致蛋白质变性后,易被蛋白酶水解,其原因是__________________ 。
(4)现有长势相同的小麦幼苗若干,完全培养液和缺Mg的完全培养液及相关器材。利用上述材料设计实验,验证Mg是小麦生长的必需元素。写出实验思路并对实验现象及结论做出分析________ 。
表一
元素 | C | O | H | N | K | Ca | Mg | P | S | Cl |
干重比例(%) | 45 | 45 | 6 | 1.5 | 1 | 0.5 | 0.2 | 0.2 | 0.1 | 0.01 |
表二
化合物 | 硝酸钾 | 硝酸钙 | 磷酸二氢氨 | 硫酸镁 | 氯化钾 | …… |
每升溶液中的体积(ml) | 6 | 4 | 2 | 1 | 2 |
表三
元素 | N | K | Ca | P | S | Mg | Cl |
元素的质量浓度(mg/L) | 224 | 235 | 160 | 62 | 32 | 24 | 1.77 |
(1)镁元素是构成
(2)与表一相比,表三中缺少的元素有
(3)小麦种子中蛋白质含量占11%左右,小麦面粉做成熟食后更容易消化吸收。从蛋白质结构分析,高温导致蛋白质变性后,易被蛋白酶水解,其原因是
(4)现有长势相同的小麦幼苗若干,完全培养液和缺Mg的完全培养液及相关器材。利用上述材料设计实验,验证Mg是小麦生长的必需元素。写出实验思路并对实验现象及结论做出分析
您最近一年使用:0次
名校
3 . 鸟的性别决定方式为ZW型。某种鸟的正常眼和疤眼是一对相对性状(基因为A、a);B、b基因位于常染色体上,bb可使部分应表现为疤眼的个体表现为正常眼,其余基因型无此影响。从群体中随机选取三对组合进行杂交实验,如下表所示。下列说法错误的是( )
杂交组合 | 子代 |
组合一:疤眼雄鸟×正常眼雌鸟 | 正常眼雄鸟:疤眼雌鸟=1∶1 |
组合二:正常眼雄鸟×疤眼雌鸟 | 全为正常眼 |
组合三:正常眼雄鸟×正常眼雌鸟 | 出现疤眼雄鸟 |
| A.疤眼由Z染色体上的a基因控制 |
| B.组合一亲本疤眼雄鸟不可能含有b基因 |
| C.组合二子代正常眼雄鸟全为杂合子 |
| D.组合三子代疤眼雄鸟可能为纯合子 |
您最近一年使用:0次
2022-03-03更新
|
809次组卷
|
5卷引用:2022届山东省潍坊市高三一模生物试题
2022届山东省潍坊市高三一模生物试题(已下线)三轮冲刺卷15-【赢在高考·黄金20卷】备战2022年高考生物模拟卷(山东专用)陕西省延安市一中2021-2022学年高一下学期第二次月考生物试题山东省枣庄市滕州一中2022-2023学年高三10月过关考试生物试题山东省泰安市长城中学2022-2023学年高三上学期期末生物试题
名校
4 . 溶菌酶是一种由动物细胞合成并分泌的单肽链蛋白质,它能溶解细菌的细胞壁从而使细菌解体。动物不同部位分泌的溶菌酶在结构上存在一定差异,研究人员比较了肾溶菌酶和胃溶菌酶的氨基酸组成,结果如下表:
注:“+”表示是此氨基酸,“-”表示否
(1)氨基酸通过_______ (结合方式)形成溶菌酶,这些氨基酸形成溶菌酶的过程中,生成_____ 分子的水。根据表中数据,仅从氨基酸的角度分析,肾溶菌酶和胃溶菌酶在功能上存在差异的原因是______ 。
(2)科学家通过3H标记的亮氨酸探究溶菌酶合成和分泌的过程,该种方法称为________ 。溶菌酶首先在_______ 开始多肽链的合成,细胞中可为此过程提供能量的场所有_______ 。
(3)科学家研究发现,面向内质网一侧的高尔基体膜成分与内质网膜成分类似,面向细胞膜一侧的高尔基体膜成分与细胞膜成分类似。由该事实推测高尔基体在溶菌酶的合成和分泌过程中的作用是________ 。
(4)溶菌酶可被用于细菌引起的疾病的治疗,但是对具细胞壁的真菌引起的疾病不发挥作用,原因是________ 。
| 氨基酸数目 | 第80位天冬酰胺氨基酸 | 第75位天冬氨酸 | 第50位谷氨酸 | 精氨酸数目 | |
| 肾溶菌酶 | 130 | - | - | - | 3 |
| 胃溶菌酶 | 130 | + | + | + | 8 |
注:“+”表示是此氨基酸,“-”表示否
(1)氨基酸通过
(2)科学家通过3H标记的亮氨酸探究溶菌酶合成和分泌的过程,该种方法称为
(3)科学家研究发现,面向内质网一侧的高尔基体膜成分与内质网膜成分类似,面向细胞膜一侧的高尔基体膜成分与细胞膜成分类似。由该事实推测高尔基体在溶菌酶的合成和分泌过程中的作用是
(4)溶菌酶可被用于细菌引起的疾病的治疗,但是对具细胞壁的真菌引起的疾病不发挥作用,原因是
您最近一年使用:0次
2023-02-13更新
|
85次组卷
|
2卷引用:山东省潍坊市2022-2023学年高一下学期开学测试生物试题
5 . 果蝇是遗传学研究的良好材料,其特点有∶①生活史短,从初生卵发育至新羽化的成虫大约为10~12 天,成虫存活大约15 天;②性别决定方式为XY型,具体的决定方式如表1所示,其中在性染色体组成为XXY雌果蝇中,XY联会的概率远低于 XX 联会,另外雌果蝇的结构很特别,有一个储精囊,交配后便会将雄果蝇的精子储存在储精囊中,之后这只雌果蝇便能不断地产生此次交配的子代;③突变型多,已知果蝇的部分突变型如表2所示。摩尔根及其学生利用果蝇作为实验材料,通过实验证明了基因位于染色体上,并给出了第一幅果蝇多种基因在染色体上的相对位置图。
表1
表2
(1)摩尔根利用在一群红眼果蝇中发现的一只白眼雄果蝇,做了著名的“摩尔根果蝇杂交实验”,即用红眼雌果蝇与该白眼雄果蝇杂交得F1,再让 F1自由交配得F2.基于F2的实验结果摩尔根提出了______ 的“假说”,并预测了测交的实验结果。请基于"摩尔根果蝇杂交实验",结合果蝇的特点完成后续测交实验的设计思路______ 。
(2)摩尔根的学生重复做了红眼雄果蝇与白眼雌果蝇的杂交实验,发现子代 2000 ~3000 只红眼雌果蝇中会出现一只“白眼雌果蝇”,同时又在 2000~3000只“白眼雄果蝇”中会出现一只红眼雄果蝇。
①某生物兴趣小组一致认为该现象不是基因突变导致的,推测持该观点的理由是______ 。
②显微镜观察发现“白眼雌果蝇”有三条性染色体,则进一步将其和野生型的红眼雄果蝇进行交配,推测其子代情况应为______ 。
(3)现有三种不同品系(纯合体)的果蝇,1号为红眼长翅直刚毛果蝇;2号为残翅果蝇;3号为白眼小翅卷刚毛果蝇(不考虑性染色体组成异常)。
①果蝇的翅型有3种类型∶长翅、小翅和残翅,由两对等位基因共同决定,当个体中Vg和 M基因同时存在时,表现为长翅,Vg基因不存在时,表现为残翅,其余表现为小翅。某生物兴趣小组随机选取1号与2号的两只果蝇杂交得F1,F1出现小翅型,则 F1雌雄果蝇交配得F2的翅型及比例为___ 。
②为确定基因在染色体上的相对位置,摩尔根和他的学生们依据染色体上两基因距离越远,它们之间的染色体互换的机会就越多,反之越少的现象发明了三点测交法,即利用三杂合体与三隐性纯合体做测交实验。对同一染色体上的三种基因来说,染色体的互换可归为三类,即一种两种单交换型和双交换型,如下图所示。现为确定w、sn、m 三种基因的位置关系,某生物兴趣小组选用1号的雄果蝇和3号雌果蝇做杂交得F1,再用F1的雌果蝇与3号的雄果蝇做测交实验,子代的表型及数量如下表所示。
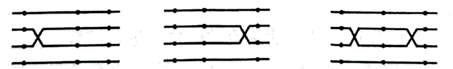
分析表中结果可知,F1的雌果蝇产生的配子有___ 种,w、sn、m 三种基因在X染色体上的顺序为______ 。
表1
| 性染色体组成 | 性别 |
| XX、XXY | 雌性 |
| XY、XYY、XO | 雄性 |
| XXX、YO、YY | 致死 |
| 影响部分 | 性状表现 | 基因符号 | 所在染色体 |
| 翅型 | 长翅、残翅 | Vg、vg | Ⅱ |
| 眼色 | 红眼、白眼 | W、w | X |
| 刚毛 | 直刚毛、卷刚毛 | Sn、sn | X |
| 翅型 | 长翅、小翅 | M、m | X |
(2)摩尔根的学生重复做了红眼雄果蝇与白眼雌果蝇的杂交实验,发现子代 2000 ~3000 只红眼雌果蝇中会出现一只“白眼雌果蝇”,同时又在 2000~3000只“白眼雄果蝇”中会出现一只红眼雄果蝇。
①某生物兴趣小组一致认为该现象不是基因突变导致的,推测持该观点的理由是
②显微镜观察发现“白眼雌果蝇”有三条性染色体,则进一步将其和野生型的红眼雄果蝇进行交配,推测其子代情况应为
(3)现有三种不同品系(纯合体)的果蝇,1号为红眼长翅直刚毛果蝇;2号为残翅果蝇;3号为白眼小翅卷刚毛果蝇(不考虑性染色体组成异常)。
①果蝇的翅型有3种类型∶长翅、小翅和残翅,由两对等位基因共同决定,当个体中Vg和 M基因同时存在时,表现为长翅,Vg基因不存在时,表现为残翅,其余表现为小翅。某生物兴趣小组随机选取1号与2号的两只果蝇杂交得F1,F1出现小翅型,则 F1雌雄果蝇交配得F2的翅型及比例为
②为确定基因在染色体上的相对位置,摩尔根和他的学生们依据染色体上两基因距离越远,它们之间的染色体互换的机会就越多,反之越少的现象发明了三点测交法,即利用三杂合体与三隐性纯合体做测交实验。对同一染色体上的三种基因来说,染色体的互换可归为三类,即一种两种单交换型和双交换型,如下图所示。现为确定w、sn、m 三种基因的位置关系,某生物兴趣小组选用1号的雄果蝇和3号雌果蝇做杂交得F1,再用F1的雌果蝇与3号的雄果蝇做测交实验,子代的表型及数量如下表所示。
| 表型 | 红长直 | 白小卷 | 白小直 | 红长卷 | 红小直 | 白长卷 | 白长直 | 红小卷 |
| 数量 | 520 | 490 | 5 | 4 | 37 | 36 | 24 | 25 |
分析表中结果可知,F1的雌果蝇产生的配子有
您最近一年使用:0次
2022-04-28更新
|
1228次组卷
|
2卷引用:2022届山东省潍坊市高三二模生物试题
名校
解题方法
6 . 科研人员为探究Mg2+对水稻光合作用的影响,开展了一系列实验。请回答:
(1)Mg2+参与光合作用过程中____ (物质)的合成,该物质直接参与的反应阶段可为暗反应____ 中(物质变化)提供能量物质____ 。
(2)为研究Mg2+对光合作用的影响,科研人员分别模拟环境中Mg2+正常供给( + Mg2+)、缺乏(-Mg2+)条件,测定水稻光合作用相关指标,如图1、2所示。____ ”的节律性波动,且Mg2+可以显著____ 白天固定CO2的过程。
②进一步测定上述过程中酶R(催化C5与CO2的反应)的变化如图2,结果表明Mg2+很可能通过____ ,从而促进CO2的固定。
(3)为探究叶绿体中Mg2+节律性波动的原因,科研人员又对多种突变体水稻进行实验。____ 。
②在另一株突变体OS(OS基因缺失)中,白天叶绿体中Mg2+含量显著升高。据此,对MT3蛋白、OS蛋白的作用关系,科研人员提出如下假设:
假设1:OS蛋白抑制MT3蛋白,并调节其节律性运输Mg2+至叶绿体内。
假设2:MT3蛋白节律性运输Mg2+至叶绿体内,而OS蛋白运出Mg2+。
通过检测野生型和多个突变体的Mg2+含量,如下表。
(【注】:双突变体OM指OS基因和MT3基因均缺失,且实验中不考虑Mg2+的损耗)
若表中①为____ (填字母),则说明假设一是正确的。
A.1.5 B.2.5 C.3 D.3.5
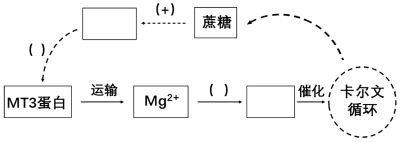
(4)已有研究表明,光合作用产生的蔗糖会影响OS蛋白的相对含量,且对光合作用进行负反馈调节。结合本实验研究,完善下列白天水稻叶绿体中Mg2+调节光合作用及其节律性变化的模型[方框中填写物质名称,( )中选填“+”表示促进、“-”表示抑制]。
(1)Mg2+参与光合作用过程中
(2)为研究Mg2+对光合作用的影响,科研人员分别模拟环境中Mg2+正常供给( + Mg2+)、缺乏(-Mg2+)条件,测定水稻光合作用相关指标,如图1、2所示。
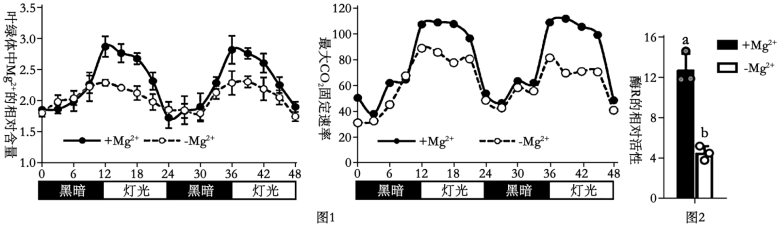
②进一步测定上述过程中酶R(催化C5与CO2的反应)的变化如图2,结果表明Mg2+很可能通过
(3)为探究叶绿体中Mg2+节律性波动的原因,科研人员又对多种突变体水稻进行实验。
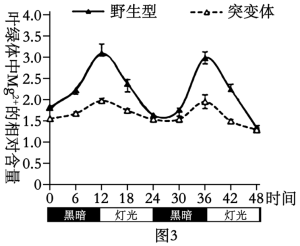
②在另一株突变体OS(OS基因缺失)中,白天叶绿体中Mg2+含量显著升高。据此,对MT3蛋白、OS蛋白的作用关系,科研人员提出如下假设:
假设1:OS蛋白抑制MT3蛋白,并调节其节律性运输Mg2+至叶绿体内。
假设2:MT3蛋白节律性运输Mg2+至叶绿体内,而OS蛋白运出Mg2+。
通过检测野生型和多个突变体的Mg2+含量,如下表。
| 序号 | 水稻植株 | 叶绿体中Mg2+相对含量 |
| 1 | 野生型 | 2.5 |
| 2 | 突变体MT3 | 1.5 |
| 3 | 突变体OS | 3.5 |
| 4 | 双突变体OM | ① |
若表中①为
A.1.5 B.2.5 C.3 D.3.5
(4)已有研究表明,光合作用产生的蔗糖会影响OS蛋白的相对含量,且对光合作用进行负反馈调节。结合本实验研究,完善下列白天水稻叶绿体中Mg2+调节光合作用及其节律性变化的模型[方框中填写物质名称,( )中选填“+”表示促进、“-”表示抑制]。
您最近一年使用:0次
2024-05-10更新
|
549次组卷
|
5卷引用:山东省潍坊实验中学2023-2024学年高三下学期5月份三模拉练试题
名校
解题方法
7 . 棉花(二倍体)植株中花序分枝有关基因分别位于1号和3号染色体上。利用花序分枝过多纯合体甲进行诱变育种,获得分枝不增加的纯合体乙,甲与乙杂交获得F1,F1自交,F2表现为不同程度的分枝,对F2部分个体的花序分枝相关基因进行测序并统计花序分枝数,结果如下表所示。研究发现,突变体乙的3号染色体上花序分枝有关基因和甲的种类相同,但数量不同。下列说法正确的是( )
注:“+”表示相关DNA序列与甲中的一致,“-”表示相关 DNA序列与甲的不一致
F2 | 花序分枝数(个) | 1号染色体相关序列 | 3号染色体相关序列 |
1 | 20 | +/+ | +/+ |
2 | 16 | +/- | +/+ |
3 | 13 | -/- | +/+ |
4 | 6 | +/+ | +/- |
5 | 4 | +/+ | -/- |
6 | 3 | -/- | +/- |
7 | 1 | -/- | -/- |
| A.花序分枝数取决于花序分枝有关基因的种类和数量 |
| B.3号染色体上花序分枝有关基因对花序分枝数的影响更大 |
| C.F2中表型与F1相同的植株占比约为3/8 |
| D.突变体乙的3号染色体结构发生改变 |
您最近一年使用:0次
2024-03-11更新
|
757次组卷
|
7卷引用:山东省潍坊实验中学2023-2024学年高三下学期5月份三模拉练试题
名校
8 . 植物生命活动受植物激素的调控。下图一表示种子在解除休眠过程中几种激素的变化情况;图二表示生长素浓度对黄豆根和茎生长的影响;表是不同浓度的生长素溶液对玫瑰插条生根影响的实验结果。请分析回答:
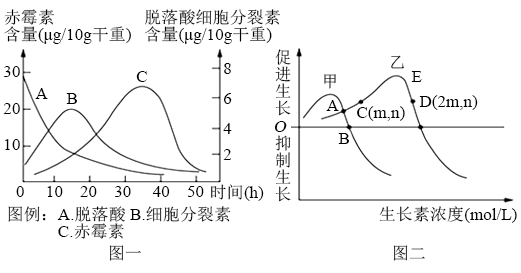
(1)生长素的化学本质是____________ ,生长素促进植物生长的实质是_________________________ 。
(2)水平放置的黄豆幼苗根会表现为向地弯曲生长,原因是______________________________ 。如果黄豆幼苗已经表现出向光性,通过一定方法测得黄豆幼苗向光面的生长素浓度为m,则其背光面生长素浓度为_____________ 。
(3)种子在解除休眠过程中,脱落酸和赤霉素表现为___________ 作用,图一的结果说明__________________________________ 。
(4)请在表三的基础上进一步完成探究生长素促进攻瑰插条生根的最适浓度范围的实验步骤:
①在浓度_____________________________ 的生长素溶液9组。
②_______________________________ ,并编号。
③分别用_________________________ 的上述配制的生长素溶液处理对应组的玫瑰插条相同时间。
④在相同且适宜条件下培养一-段时间,测量并记录玫瑰插条的根长度,并________________________ 。
编号 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
生长素溶液浓度(mg/mL) | 0(清水) | 0.2 | 0.4 | 0.6 | 0.8 | 1.0 | 1.2 |
平均根长(cm) | 2.2 | 2.8 | 3.6 | 5.0 | 5.0 | 4.2 | 3.1 |
(2)水平放置的黄豆幼苗根会表现为向地弯曲生长,原因是
(3)种子在解除休眠过程中,脱落酸和赤霉素表现为
(4)请在表三的基础上进一步完成探究生长素促进攻瑰插条生根的最适浓度范围的实验步骤:
①在浓度
②
③分别用
④在相同且适宜条件下培养一-段时间,测量并记录玫瑰插条的根长度,并
您最近一年使用:0次
名校
9 . 下图中地雀A与地雀B可以交配产生可育后代,地雀C与地雀A、B之间不能互相交配。图1、2、3是关于地雀的进化、发展过程。
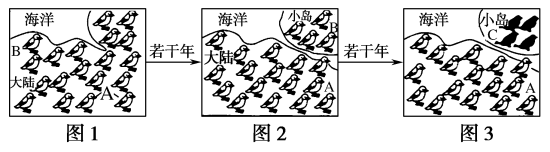
(1)上述地雀之间的关系能体现遗传多样性的类型是________________ ,能体现物种多样性的类型是____________________________ 。
(2)若干年后,小岛上出现了地雀C.在这一变化过程中,起到选择作用的主要因素是_________________ 。这种进化的实质上是由于不同种群的______________ 发生改变。
(3)若图2时期,小岛和大陆的两个地雀种群的相关基因型种类和个体数如下表所示。
①种群甲的全部个体所含的全部基因,是这个种群的____________ 。
②种群甲中A基因的频率为________ 。
③已知表中种群乙的每个个体有2×104个基因,每个基因的突变概率都是10-5,则该种群中出现突变的基因数是________ 。
(1)上述地雀之间的关系能体现遗传多样性的类型是
(2)若干年后,小岛上出现了地雀C.在这一变化过程中,起到选择作用的主要因素是
(3)若图2时期,小岛和大陆的两个地雀种群的相关基因型种类和个体数如下表所示。
| 甲 | 基因数 | AATT | AaTT | aaTT | AATt | AaTt | aaTt | AAtt | Aatt | aatt | 总数 |
| 个体数 | 20 | 22 | 4 | 20 | 12 | 3 | 9 | 8 | 2 | 100 | |
| 乙 | 基因型 | AATT | AaTT | aaTT | |||||||
| 个体数 | 196 | 168 | 36 | 400 |
①种群甲的全部个体所含的全部基因,是这个种群的
②种群甲中A基因的频率为
③已知表中种群乙的每个个体有2×104个基因,每个基因的突变概率都是10-5,则该种群中出现突变的基因数是
您最近一年使用:0次
2021-12-20更新
|
78次组卷
|
10卷引用:山东省寿光市第一中学2017-2018学年高二10月月段检测生物试题
山东省寿光市第一中学2017-2018学年高二10月月段检测生物试题云南省临沧一中2017-2018学年高二上学期第一次月考理综生物试题【全国百强校】江西省南昌市第十中学2019届高三上学期第二次月考生物试题(已下线)2019年高考二轮复习综合强化—专题11 现代生物进化理论(已下线)专题7.5 现代生物进化理论(练)——《2020年高考一轮复习讲练测》广东省实验中学2020-2021学年高二上学期期中生物试题福建省三明市教研联盟校2021-2022学年高二上学期期中联考生物试题云南省临沧一中2017-2018学年高二(上)第一次月考生物试题陕西省咸阳市实验中学2022-2023学年高一下学期第三次月考生物试题海南省文昌市文昌中学2023-2024学年高三上学期期末生物试题B卷
名校
解题方法
10 . 科研人员为研究遮阴条件对黄瓜苗生长发育和光合作用的影响,通过盆栽实验,测定了75%遮阴处理和自然条件下黄瓜苗的相关生理指标,实验结果如下表。
(1)通常采用_________ (试剂)对黄瓜苗绿叶中光合色素进行提取。在新鲜叶片量一定的情况下,为了提高提取液色素浓度,可以采取的措施有_________ (答出2点即可)。
(2)75%遮阴处理后,黄瓜苗叶绿素含量增加,这对植物适应弱光环境的意义是_________ 。黄瓜苗产生的适应弱光环境的变化还有_________ 。
(3)据表分析,与自然条件下相比,75%遮阴处理胞间CO2浓度较高的原因是_________ 。
(4)研究表明,弱光能间接影响光合产物在植物各器官间的运输和分配。由此可推测,75%遮阴处理黄瓜苗的平均株高高于自然条件下的原因是_________ 。
叶面积(cm2) | 总叶绿素(mg·g-1FW) | 胞间CO2浓度(μmol·mol-1) | 净光合速率(μmol·m-2·s-1) | |
自然条件 | 2856 | 1.41 | 178 | 14.05 |
75%遮阴 | 3715 | 1.67 | 297 | 4.47 |
(2)75%遮阴处理后,黄瓜苗叶绿素含量增加,这对植物适应弱光环境的意义是
(3)据表分析,与自然条件下相比,75%遮阴处理胞间CO2浓度较高的原因是
(4)研究表明,弱光能间接影响光合产物在植物各器官间的运输和分配。由此可推测,75%遮阴处理黄瓜苗的平均株高高于自然条件下的原因是
您最近一年使用:0次
2024-05-24更新
|
181次组卷
|
2卷引用:山东省潍坊市2023—2024学年四县联考高三下学期三模考试生物试题


