名校
1 . 早在一百多年前,果蝇就走进了科学家的实验室,成为实验室里的明星模式生物,1907年摩尔根在哥伦比亚大学建立了著名的蝇室,形成了一个大的果蝇遗传学研究中心。
(1)果蝇作为遗传学研究中的模式生物,其优点有______ (至少答两点)。
(2)摩尔根利用果蝇做了如下图所示杂交实验,此外他还将F1红眼雌果蝇与亲代白眼雄果蝇进行交配,发现后代中白眼雌果蝇占______ ,该交配方式称为______ 。______ 进行交配,若假说成立,那么子代将出现雌果蝇都是红眼,雄果蝇都是白眼。最后再通过实验来检验实验结果与演绎是否相符,这种研究方法就叫作______ 。
(4)某果蝇种群中灰体(D)对黑檀体(d)为显性,灰体纯合果蝇与黑檀体果蝇杂交,后代中出现一只黑檀体果蝇,原因可能是亲本果蝇产生配子过程中发生了基因突变或染色体片段缺失。已知一对同源染色体都缺失相同片段胚胎致死,各型配子活力相同。请完成下列实验步骤及结果预测。
实验步骤:用该黑檀体果蝇与基因型为Dd的果蝇杂交,获得F1;F1自由交配,观察、统计F2的表现型及比例。
结果预测:a、如果F2表现型及比例为灰体:黑檀体=______ ,则为基因突变;b、如果F2表现型及比例为灰体:黑檀体=______ ,则为染色体片段缺失。
(5)该果蝇种群中眼色有四种,M基因控制棕色素合成,N基因控制朱红色素合成。棕色素和朱红色素同时存在时表现暗红眼,两种色素都不存在时表现白眼。现将一只白眼果蝇与纯合暗红眼果蝇杂交,F1全部为暗红眼,F1与白眼果蝇杂交实验结果如下表所示:
根据表中数据推测F1作______ (父本/母本)时发生了互换,F1的M基因与N基因______ (在/不在)同一条染色体上,杂交组合Ⅱ中,F1产生的雌配子的类型及其比例是______ 。
(1)果蝇作为遗传学研究中的模式生物,其优点有
(2)摩尔根利用果蝇做了如下图所示杂交实验,此外他还将F1红眼雌果蝇与亲代白眼雄果蝇进行交配,发现后代中白眼雌果蝇占

(4)某果蝇种群中灰体(D)对黑檀体(d)为显性,灰体纯合果蝇与黑檀体果蝇杂交,后代中出现一只黑檀体果蝇,原因可能是亲本果蝇产生配子过程中发生了基因突变或染色体片段缺失。已知一对同源染色体都缺失相同片段胚胎致死,各型配子活力相同。请完成下列实验步骤及结果预测。
实验步骤:用该黑檀体果蝇与基因型为Dd的果蝇杂交,获得F1;F1自由交配,观察、统计F2的表现型及比例。
结果预测:a、如果F2表现型及比例为灰体:黑檀体=
(5)该果蝇种群中眼色有四种,M基因控制棕色素合成,N基因控制朱红色素合成。棕色素和朱红色素同时存在时表现暗红眼,两种色素都不存在时表现白眼。现将一只白眼果蝇与纯合暗红眼果蝇杂交,F1全部为暗红眼,F1与白眼果蝇杂交实验结果如下表所示:
| 杂交组合 | 父本 | 母本 | F2表现型及比例 |
| Ⅰ | F1 | 白眼 | 暗红眼:白眼=1:1 |
| Ⅱ | 白眼 | F1 | 暗红眼:棕眼:朱红眼:白眼=43:7:7:43 |
您最近一年使用:0次
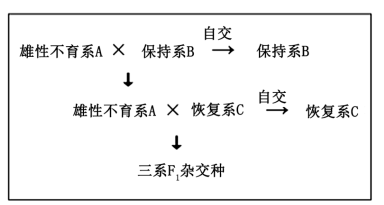
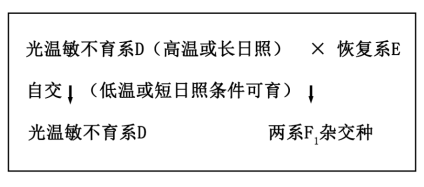
2 . 水稻(2n=24)花为两性花,一株稻穗约200~300朵小花,花粉自然条件下存活不足5分钟。袁隆平院士及其团队研发的三系杂交水稻和两系杂交水稻,为我国粮食安全作出巨大贡献。以下是两种杂交育种过程模式图及三系杂交育种体系中与雄性不育有关基因分布表,细胞核基因R1对r1、R2对r2均表现显性,且仅有基因型S(r1r1r2r2)的水稻表现为雄性不育,请回答:
三系杂交水稻,育种体系由保持系、雄性不育系和恢复系组成,保持系和恢复系能够通过自交进行自身品系的保持,而雄性不育系通过与保持系杂交维持雄性不育品系,雄性不育系与恢复系杂交获得杂交种F1;两系杂交水稻,育种体系只需雄性不育系和恢复系,雄性不育系在短日照或低温条件下可育,雄性不育系在长日照或高温条件下与恢复系杂交获得杂交种F1。
(1)水稻无法通过人工去雄的方法大量生产杂交种的原因是_________ 。
(2)三系杂交育种体系中,保持系基因型为_________ 。
(3)为研究水稻雄性不育的遗传规律,科研人员用基因型为S(r1r1r2r2)和N(R1R1R2R2)的亲本杂交得到F1,F1自交,F2统计结果如下表。回答下列问题:(注:雄性不育株结实率为0,雄性可育株结实率大于0)
①据表中数据分析可知R1(r1)和R2(r2)两对等位基因遵循自由组合定律,理由是_________ 。
②写出F2中30%≤f<70%的植株的所有基因型_________ 。
(4)两系杂交中光温敏水稻在不同条件下育性不同的根本原因是_________ 。与三系杂交相比,两系杂交育种的优点是_________ 。
三系杂交水稻,育种体系由保持系、雄性不育系和恢复系组成,保持系和恢复系能够通过自交进行自身品系的保持,而雄性不育系通过与保持系杂交维持雄性不育品系,雄性不育系与恢复系杂交获得杂交种F1;两系杂交水稻,育种体系只需雄性不育系和恢复系,雄性不育系在短日照或低温条件下可育,雄性不育系在长日照或高温条件下与恢复系杂交获得杂交种F1。
细胞质 | 细胞核 | |
雄性不育基因 | S | r1,r2 |
雄性可育基因 | N | R1,R2 |
(1)水稻无法通过人工去雄的方法大量生产杂交种的原因是
(2)三系杂交育种体系中,保持系基因型为
(3)为研究水稻雄性不育的遗传规律,科研人员用基因型为S(r1r1r2r2)和N(R1R1R2R2)的亲本杂交得到F1,F1自交,F2统计结果如下表。回答下列问题:(注:雄性不育株结实率为0,雄性可育株结实率大于0)
结实率(f)分布范围 | f=0 | 0<f<30% | 30%≤f<70% | 70%≤f<90% | 90%≤f<100% |
实测平均结实率 | 0 | 17.1% | 61.2% | 76.6% | 90.5% |
实测株数 | 3 | 16 | 22 | 14 | 2 |
子二代株数比例 | 1 | 4 | 6 | 4 | 1 |
②写出F2中30%≤f<70%的植株的所有基因型
(4)两系杂交中光温敏水稻在不同条件下育性不同的根本原因是
您最近一年使用:0次
2024-04-30更新
|
29次组卷
|
2卷引用:(教研室的资料)2024届湖南省岳阳市高三下学期三模生物试题
名校
3 . 野生型果蝇(2n=8)都是灰体红眼的纯合子,在自然繁殖过程中会出现某些性状的突变体。某兴趣小组同学以野生型及相关突变型果蝇为材料,开展有关遗传学问题的研究和分析。回答下列有关问题:
(1)野生型雄果蝇的体细胞中染色体形态有________ 种,有丝分裂后期有________ 个染色体组。
(2)果蝇的红眼和白眼由一对等位基因A/a控制。一只野生型雄果蝇与一只白眼雌果蝇杂交,F1中红眼均为雌果蝇,白眼均为雄果蝇,说明基因A/a位于________ 染色体上。F1果蝇随机交配得F2,F2雌果蝇中纯合子占________ 。
(3)已知果蝇的灰体和黑体受一对常染色体上的等位基因控制。以野生型灰体果蝇与突变型黑体果蝇为实验材料,设计一次杂交实验探究此相对性状的显隐关系,只写出实验思路即可。
实验思路:________________________________ 。
(4)果蝇翅型的野生型和长翅由一对基因B/b控制,野生型和残翅由另一对基因D/d控制。两只野生型果蝇相互交配,F1表型及比例为野生型♀∶残翅♀∶野生型♂∶长翅♂∶残翅♂∶无翅♂=6∶2∶3∶3∶1∶1。
①两对基因在染色体上的位置分别是________ ,F1的野生型雌果蝇与无翅雄果蝇相互交配,F2的野生型雌果蝇中杂合子所占比例为________ 。
②果蝇的性别与性染色体组成的关系如下表所示。
(注:性染色体组成为Y0、XXX、YY的个体均在胚胎时期死亡)
由此表可知果蝇的性别并不是由Y染色体决定,而是由________ 决定。
(1)野生型雄果蝇的体细胞中染色体形态有
(2)果蝇的红眼和白眼由一对等位基因A/a控制。一只野生型雄果蝇与一只白眼雌果蝇杂交,F1中红眼均为雌果蝇,白眼均为雄果蝇,说明基因A/a位于
(3)已知果蝇的灰体和黑体受一对常染色体上的等位基因控制。以野生型灰体果蝇与突变型黑体果蝇为实验材料,设计一次杂交实验探究此相对性状的显隐关系,只写出实验思路即可。
实验思路:
(4)果蝇翅型的野生型和长翅由一对基因B/b控制,野生型和残翅由另一对基因D/d控制。两只野生型果蝇相互交配,F1表型及比例为野生型♀∶残翅♀∶野生型♂∶长翅♂∶残翅♂∶无翅♂=6∶2∶3∶3∶1∶1。
①两对基因在染色体上的位置分别是
②果蝇的性别与性染色体组成的关系如下表所示。
性染色体组成 | XX | XY | XXY | XO | XYY |
性别 | 雌性 | 雄性,可育 | 雌性 | 雄性,不育 | 雄性,可育 |
由此表可知果蝇的性别并不是由Y染色体决定,而是由
您最近一年使用:0次
2024-04-19更新
|
199次组卷
|
5卷引用:2024届黑龙江省友谊县高级中学高三下学期第三次模拟生物试题
4 . 研究发现某二倍体作物的雄性育性可能与8号染色体的某区段的A、B、C、D等基因有关。对作物的品系甲该段染色体测序,发现与野生型植株相比,品系甲该区段均少了基因C和D,两者的花粉均可育。为寻找影响作物育性的基因,对野生型品系进行基因敲除,获得敲除纯合子(无法获得D基因敲除纯合个体),进行杂交实验结果如下表。回答下列问题:
(1)由表可知,______ 基因可导致花粉育性降低,判断依据是______ 。
(2)进一步研究发现,所有杂交组合的F1均能正常完成减数分裂。且50%的花粉可育的F1可育花粉的8号染色体都来自野生型,不育花粉的8号染色体都来自品系甲。
①推测D基因的作用是______ ,D基因发挥作用的时间是______ 。杂交组合1的F1不育配子的基因型是______ 。
②杂交组合1的F1进行自交,子代的育性及比例是______ 。
(3)为验证上述推测,向杂交组合1的F1转入1个D基因,转入之后对植物其他生理功能无影响。之后进行转基因植株的筛选和鉴定。
①筛选得到植株1,该植株花粉均可育,若上述推测正确,说明D基因转入位置在______ 染色体上。
②筛选得到植株2,该植株50%的花粉可育,若上述推测正确,说明D基因转入的位置在______ 染色体上。
③若D基因插入在5号染色体,若上述推测正确,该植株可育花粉的比例是______ 。自交子代中100%花粉可育的植株占______ 。
杂交组合 | 母本 | 父本 | F1育性性状 |
1 | 野生型 | 品系甲 | 50%的花粉可育 |
2 | A基因敲除的纯合野生品系乙 | 品系甲 | 50%的花粉可育 |
3 | B基因敲除的纯合野生品系丙 | 品系甲 | 50%的花粉可育 |
4 | C基因敲除的纯合野生品系丁 | 品系甲 | 100%的花粉可育 |
(2)进一步研究发现,所有杂交组合的F1均能正常完成减数分裂。且50%的花粉可育的F1可育花粉的8号染色体都来自野生型,不育花粉的8号染色体都来自品系甲。
①推测D基因的作用是
②杂交组合1的F1进行自交,子代的育性及比例是
(3)为验证上述推测,向杂交组合1的F1转入1个D基因,转入之后对植物其他生理功能无影响。之后进行转基因植株的筛选和鉴定。
①筛选得到植株1,该植株花粉均可育,若上述推测正确,说明D基因转入位置在
②筛选得到植株2,该植株50%的花粉可育,若上述推测正确,说明D基因转入的位置在
③若D基因插入在5号染色体,若上述推测正确,该植株可育花粉的比例是
您最近一年使用:0次
2024-03-01更新
|
271次组卷
|
3卷引用:江苏省南京市江宁区等5地2023-2024学年高三下学期开学生物试题
5 . 果蝇的长翅(B)与短翅(b)、红眼(A)与白眼(a)是两对相对性状。Y染色体上无相关基因,亲代雌果蝇与雄果蝇杂交,F1表型及数量如下表:
请回答:
(1)果蝇眼色性状的基因位于___ 染色体上,其遗传符合___ 定律,基因A与a互为___ 基因。
(2)亲本的基因型为父本___ 、母本___ 。F1长翅红眼雌果蝇的基因型有___ 种,其中杂合子:纯合子=___ 。F1长翅红眼雄果蝇的一个细胞中最多有___ 条Y染色体(不考虑变异)。
(3)现有1只长翅白眼雌果蝇与1只长翅红眼雄果蝇杂交,子代雌果蝇中长翅红眼占3/4,则子代雌果蝇中出现短翅红眼的概率为___ 。
(4)已知含1条X染色体的果蝇表现为雄性,如性染色体组成为XY,XYY,XO(不可育),含2条X染色体的果蝇表现为雌性,如性染色体组成为XX、XXY,其余性染色体组成异常的果蝇均胚胎致死。在统计数量时发现F1中有一只白眼雌果蝇:
①若该白眼雌果蝇是基因突变导致的,则该果蝇的基因型为___ 。若该白眼雌果蝇是某一亲本减数分裂过程中染色体未分离导致的,则该果蝇的基因型为___ 。发生该异常分离时的细胞名称为___ 。
②若与亲本回交产生的子代雌果蝇中___ 则为基因突变导致;
③若与亲本回交产生的子代雌果蝇中___ 则为减数分裂过程中染色体未分离导致。
| 长翅红眼 | 长翅白眼 | 短翅红眼 | 短翅白眼 | |
| 雌蝇(只) | 151 | 0 | 52 | 0 |
| 雄蝇(只) | 77 | 75 | 25 | 26 |
(1)果蝇眼色性状的基因位于
(2)亲本的基因型为父本
(3)现有1只长翅白眼雌果蝇与1只长翅红眼雄果蝇杂交,子代雌果蝇中长翅红眼占3/4,则子代雌果蝇中出现短翅红眼的概率为
(4)已知含1条X染色体的果蝇表现为雄性,如性染色体组成为XY,XYY,XO(不可育),含2条X染色体的果蝇表现为雌性,如性染色体组成为XX、XXY,其余性染色体组成异常的果蝇均胚胎致死。在统计数量时发现F1中有一只白眼雌果蝇:
①若该白眼雌果蝇是基因突变导致的,则该果蝇的基因型为
②若与亲本回交产生的子代雌果蝇中
③若与亲本回交产生的子代雌果蝇中
您最近一年使用:0次
解题方法
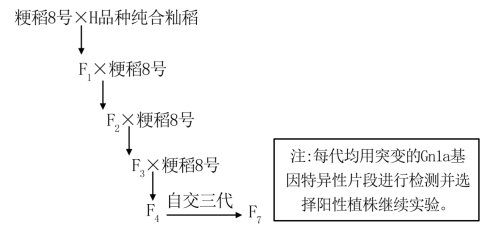
6 . 我国种植的水稻有籼稻和粳稻,籼稻广泛种植于南方,粳稻广泛种植于北方。选育籼粳杂交稻是南方水稻产业调整的重要措施。水稻单株产量主要由单株穗数、穗粒数和千粒重决定。Gnla基因(位于细胞核)通过编码细胞分裂素氧化酶,降低细胞分裂素含量,减少水稻枝梗,进而导致穗粒数减少。研究发现H品种籼稻的Gnla基因突变后能显著增加穗粒数,提高产量。目前,突变的Gnla基因广泛应用在籼稻上,但对粳稻的影响还未知,研究人员对此开展研究。研究过程如图所示,突变的Gnlα基因对粳稻8号的产量影响如下表所示。
回答下列问题
(1)H品种籼稻的Gnla基因突变后,__________ ,促进水稻枝梗数增加,从而提高穗粒数。
(2)F1-F3均和粳稻8号杂交并进行选择的目的是__________ 。检测F4时,阳性植株比例为__________ 。
F7中Gnla基因的检测结果与__________ 的检测结果完全相同,即为目标植株。
(3)研究发现,突变的Gnla基因可显著提高粳稻8号的产量。据表分析,原因是__________ 。
(4)J品种籼稻为感光型品种,粳稻8号为感温型品种,J品种籼稻和粳稻8号杂交后代的“籼(感光性)-粳(感温性)”性状会出现差异。已知感光型品种在南方生长期较短,在北方较长:感温型品种则相反。据此分析,如何根据生长期差异选育出适合在华南地区生长的“粳型”杂交稻品种?___________
组别 | 单株穗数 | 一次枝梗数 | 一次枝梗花数 | 二次枝梗数 | 二次枝梗花数 | 千粒重/g |
突变型 | 8.16 | 13.06 | 71.59 | 47.49 | 151.31 | 27.06 |
野生型 | 11.50 | 12.16 | 71.51 | 31.73 | 98.16 | 26.58 |
(1)H品种籼稻的Gnla基因突变后,
(2)F1-F3均和粳稻8号杂交并进行选择的目的是
F7中Gnla基因的检测结果与
(3)研究发现,突变的Gnla基因可显著提高粳稻8号的产量。据表分析,原因是
(4)J品种籼稻为感光型品种,粳稻8号为感温型品种,J品种籼稻和粳稻8号杂交后代的“籼(感光性)-粳(感温性)”性状会出现差异。已知感光型品种在南方生长期较短,在北方较长:感温型品种则相反。据此分析,如何根据生长期差异选育出适合在华南地区生长的“粳型”杂交稻品种?
您最近一年使用:0次
7 . 玉米是雌雄同株异花植物,利用玉米纯合雌雄同株品系M培育出雌株突变品系,该突变品系的产生原因是2号染色体上的基因T突变为t,T对t为完全显性。将抗玉米螟的基因A转入该雌株品系中获得甲、乙两株具有玉米螟抗性的植株,但由于A基因插入的位置不同,导致甲植株的株高表现正常,乙植株矮小。为研究A基因的插入位置及其产生的影响,进行了以下实验:
实验一:品系M(TT)x甲(Att)→F1中抗螟:非抗螟约为1:1;
实验二:品系M(TT)x乙(Att)→F1中抗螟矮株:非抗螟正常株高约为1:1。
(1)实验一的F1中抗螟植株的性别表现为_________ (填“雌雄同株”“雌株”或“雌雄同株和雌株”)。选取实验一的F,抗螟植株自交,若F2中抗螟雌雄同株:抗螟雌株:非抗螟雌雄同株约为__________ 则甲中转入的A基因与t基因位于同一条染色体上。
(2)若实验一的甲中转入的A基因与t基因位于同一条染色体上,则F2中抗螟雌株的基因型是_________ 并用遗传图解表示其F2中抗螟雌雄同株与抗螟雌株杂交过程_________ 。
(3)实验二中作为母本的是_________ 。选取实验二的F1抗螟矮株自交,F2中抗螟矮株雌雄同株:
抗螟矮株雌株:非抗螟正常株高雌雄同株:非抗螟正常株高雌株约为3:1:3:1,由此可知,乙中转入的A基因不位于2号染色体上,理由是__________ 。F2中抗螟矮株所占比例低于预期值,说明A基因除导致植株矮小外,还对F1的繁殖造成影响,结合实验二的结果推断这一影响最可能是________ F2抗螟矮株中T基因的频率为__________ 。
实验一:品系M(TT)x甲(Att)→F1中抗螟:非抗螟约为1:1;
实验二:品系M(TT)x乙(Att)→F1中抗螟矮株:非抗螟正常株高约为1:1。
(1)实验一的F1中抗螟植株的性别表现为
(2)若实验一的甲中转入的A基因与t基因位于同一条染色体上,则F2中抗螟雌株的基因型是
(3)实验二中作为母本的是
抗螟矮株雌株:非抗螟正常株高雌雄同株:非抗螟正常株高雌株约为3:1:3:1,由此可知,乙中转入的A基因不位于2号染色体上,理由是
您最近一年使用:0次
名校
解题方法
8 . 棉花(二倍体)植株中花序分枝有关基因分别位于1号和3号染色体上。利用花序分枝过多纯合体甲进行诱变育种,获得分枝不增加的纯合体乙,甲与乙杂交获得F1,F1自交,F2表现为不同程度的分枝,对F2部分个体的花序分枝相关基因进行测序并统计花序分枝数,结果如下表所示。研究发现,突变体乙的3号染色体上花序分枝有关基因和甲的种类相同,但数量不同。下列说法正确的是( )
注:“+”表示相关DNA序列与甲中的一致,“-”表示相关 DNA序列与甲的不一致
F2 | 花序分枝数(个) | 1号染色体相关序列 | 3号染色体相关序列 |
1 | 20 | +/+ | +/+ |
2 | 16 | +/- | +/+ |
3 | 13 | -/- | +/+ |
4 | 6 | +/+ | +/- |
5 | 4 | +/+ | -/- |
6 | 3 | -/- | +/- |
7 | 1 | -/- | -/- |
| A.花序分枝数取决于花序分枝有关基因的种类和数量 |
| B.3号染色体上花序分枝有关基因对花序分枝数的影响更大 |
| C.F2中表型与F1相同的植株占比约为3/8 |
| D.突变体乙的3号染色体结构发生改变 |
您最近一年使用:0次
2024-03-11更新
|
760次组卷
|
7卷引用:2024届山东省烟台市高三一模生物试题
名校
解题方法
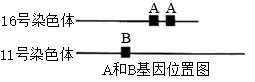
9 . 成年人的血红蛋白由两条α肽链和两条β肽链构成。在16号染色体短臂上有两个能独立控制α肽链合成的基因A,在 11号染色体上有一个控制β肽链合成的基因B,如下图所示。A基因突变导致α肽链异常,B基因突变导致β肽链异常。当患者细胞内异常肽链和正常肽链同时存在时,可随机结合形成血红蛋白,有异常肽链参与构成的血红蛋白功能异常。A 基因或B基因突变分别引起α型地中海贫血(简称α-地贫)或β型地中海贫血(简称β-地贫)。A基因个数与表型的关系如下表,回答以下问题(不考虑交叉互换)。
(1)由题干可知,16号染色体属于____ (填“端”“近端”或“中间”)着丝粒染色体。
(2)李华是一名中型α-地贫患者,他的爸爸是重型α-地贫患者。据了解,李华有一个因α-地贫胎儿期流产未出世的姐姐。据此推测,李华妈妈的表型可能有____ 种。为确认妈妈的基因型,李华带妈妈去做了基因检测,检测时应从血液样品的____ (填“红细胞”或“白细胞”)中提取 DNA。结果显示,妈妈含有2个A基因。若李华的父母还想再生育一个孩子,生出患中型α-地贫女儿的概率为____ 。若李华与一个同样患中型α-地贫的女性婚配,则其出生的后代表型可能有____ 种。
(3)某患者为β-地贫杂合体(A基因未发生突变),他体内的血红蛋白有____ 种。异常β肽链和正常β肽链相比,只有第6位谷氨酸(密码子为GAA、GAG)被缬氨酸(GUU、GUG、GUA、GUC)替代,由此推测 B 基因发生了碱基对的____ , 为确定突变基因的序列, 建议检测____ 。
A. 该突变基因的碱基组成 B. 该突变基因 mRNA 的碱基序列
C. β肽链的氨基酸序列 D. 细胞中 mRNA 的表达量
A基因数量(个) | 4 | 3 | 2 | 1 | 0 |
表型 | 正常 | 轻型α-地贫 | 中型α-地贫 | 重型α-地贫 | 胎儿期流产 |
(1)由题干可知,16号染色体属于
(2)李华是一名中型α-地贫患者,他的爸爸是重型α-地贫患者。据了解,李华有一个因α-地贫胎儿期流产未出世的姐姐。据此推测,李华妈妈的表型可能有
(3)某患者为β-地贫杂合体(A基因未发生突变),他体内的血红蛋白有
A. 该突变基因的碱基组成 B. 该突变基因 mRNA 的碱基序列
C. β肽链的氨基酸序列 D. 细胞中 mRNA 的表达量
您最近一年使用:0次
10 . 玉米是世界上最重要的粮食作物之一,南美洲和北美洲的土著人培育出了具有各种不同颜色的玉米籽粒的品种一黄色、白色、紫色等,并且赋予每种颜色不同的审美和宗教价值。例如,一些部落认为有斑点的玉米籽粒是力量和活力的象征。玉米籽粒的颜色如褐色、斑点等的遗传是不稳定的。
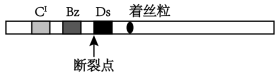
(1)种子中的胚乳是由两个极核与一个精子极核形成的。玉米籽粒的颜色与胚乳最外层糊粉层的颜色有关。极核的形成过程如下:___________ 种。
(2)玉米的第9号染色体上有两对与糊粉层颜色有关的等位基因,CI对C呈显性、Bz对bz呈显性,其中Bz促进糊粉层发育为紫色,bzbz纯合子则促进糊粉层发育为褐色,但CI抑制糊粉层颜色的发生,糊粉层表现为无色。麦克林托克选择CCbzbz的母本与CICIBzBz的父本进行杂交,得到F1代。
①理论上F1代籽粒的颜色为___________ 色。
②实际的杂交结果中,大部分籽粒的确如此,但同时还发现有一些籽粒在此颜色的背景上出现了褐色斑点。麦克林托克猜想这些杂合体中,在胚乳发育的某个时期CI基因连同___________ 一起发生了丢失,导致有些细胞群能够制造褐色色素:如果在胚乳发育早期就发生了丢失,褐色色斑的面积会___________ (“较大”、“较小”)。
③麦克林托克提出了关于有关基因丢失机制的解释:9号染色体还有一个重要的基因Ds,它是染色体发生断裂的位点,断裂后的CI所在染色体片段丢失的原因是__________ 。__________ ,在此过程中细胞发生了__________ 变异。
(3)麦克林托克随后又发现单独的Ds并不能直接诱导染色体断裂,而是需要另一个因子的帮助,即Ac。Ac位于另一条染色体上,只有在Ac存在的前体条件下,Ds才能发生断裂。更深入的研究发生,Ds不仅能诱导染色体断裂,而且能够从染色体的一个位置上切离,转移到另一个位置上。
①Ds从染色体的一个位置转移到另一个位置上,涉及____________ 键的水解与重建。
②基因型为_____________ 精子与极核结合形成为基因型CCCBzbzbzDs的胚乳,其糊粉层发育为全褐色,可能的原因是_____________ 。
(4)麦克林托克用Ac/Ds双因子系统(转座元件)解释了玉米9号染色体的遗传不稳定性。在玉米中转座元件是普遍存在的,转座元件从一个位置移动到另一个位置时,它们会造成染色体结构变异或引起____________ ,因此这些元件有着非常重要的遗传意义。
(1)种子中的胚乳是由两个极核与一个精子极核形成的。玉米籽粒的颜色与胚乳最外层糊粉层的颜色有关。极核的形成过程如下:

(2)玉米的第9号染色体上有两对与糊粉层颜色有关的等位基因,CI对C呈显性、Bz对bz呈显性,其中Bz促进糊粉层发育为紫色,bzbz纯合子则促进糊粉层发育为褐色,但CI抑制糊粉层颜色的发生,糊粉层表现为无色。麦克林托克选择CCbzbz的母本与CICIBzBz的父本进行杂交,得到F1代。
①理论上F1代籽粒的颜色为
②实际的杂交结果中,大部分籽粒的确如此,但同时还发现有一些籽粒在此颜色的背景上出现了褐色斑点。麦克林托克猜想这些杂合体中,在胚乳发育的某个时期CI基因连同
③麦克林托克提出了关于有关基因丢失机制的解释:9号染色体还有一个重要的基因Ds,它是染色体发生断裂的位点,断裂后的CI所在染色体片段丢失的原因是
(3)麦克林托克随后又发现单独的Ds并不能直接诱导染色体断裂,而是需要另一个因子的帮助,即Ac。Ac位于另一条染色体上,只有在Ac存在的前体条件下,Ds才能发生断裂。更深入的研究发生,Ds不仅能诱导染色体断裂,而且能够从染色体的一个位置上切离,转移到另一个位置上。
①Ds从染色体的一个位置转移到另一个位置上,涉及
②基因型为
(4)麦克林托克用Ac/Ds双因子系统(转座元件)解释了玉米9号染色体的遗传不稳定性。在玉米中转座元件是普遍存在的,转座元件从一个位置移动到另一个位置时,它们会造成染色体结构变异或引起
您最近一年使用:0次


