(1)请根据图片或文字选出相应的变异类型

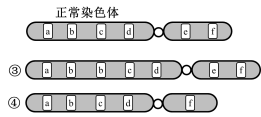

②对应的变异类型是
③对应的变异类型是
④对应的变异类型是
⑤对应的变异类型是
A.倒位 B.易位 C.缺失 D.重复
(2)在人的HD基因中,存在一段以三个相邻核酸(CAG)为单位的重复序列。亨廷顿舞蹈症患者的HD基因中,CAG的重复序列异常扩增且重复次数越多,病情越严重。由此推测导致该病的变异类型是( )
| A.基因突变 | B.染色体非整倍体变异 |
| C.基因重复 | D.染色体整倍体变异 |
一种兔的毛色由常染色体上的一对等位基因A、a控制。下表是三组亲本组合的遗传实验结果(注意:每一组的亲本都不只一对),请分析回答问题:
亲本组合 | 第一组 | 第二组 | 第三组 | |
双亲均为白毛 | 双亲中一方为白毛,另一方为黑毛 | 双亲均为黑毛 | ||
子代表现型和数量 | 黑毛 | 0 | 596 | 742 |
白毛 | 869 | 185 | 68 | |
(3)根据表中第
(4)第二组亲本中白毛兔的基因型是
①aa ②AA ③Aa
| 杂交组合 | 亲本表型 | F1表型 | F2表型及比例 |
组合一 | 红粒×白粒 | 籽粒全部含有红色素 | 含有红色素籽粒∶白粒=3∶1 |
组合二 | 红粒×白粒 | 籽粒全部含有红色素 | 含有红色素籽粒∶白粒=15∶1 |
组合三 | 红粒×白粒 | 籽粒全部含有红色素 | 含有红色素籽粒∶白粒=63∶1 |
(1)小麦籽粒颜色至少是由
(2)组合二F1的基因型为
(3)将组合一F1和组合三F1杂交,其后代白粒小麦所占比例为
(4)研究发现,基因Tamyb10能够通过激活类黄酮与脱落酸合成相关基因来调控籽粒颜色和穗发芽抗性等。经长期驯化的大多数白粒小麦品种的种皮红色素不能正常积累,与该基因突变有关。例如基因Tamyb10-A1因其有2.2kb大片段T-DNA插入而失活,该突变属于
(5)综上分析推知,基因与性状并不是简单的一一对应关系,表现在
| 项目 | 亲本 | F1表型及比例 |
| 实验一 | 子叶深绿不抗病(♀)×子叶浅绿抗病(♂) | 子叶深绿抗病:子叶浅绿抗病=1:1 |
| 实验二 | 子叶深绿不抗病(♀)×子叶浅绿抗病(♂) | 子叶深绿抗病:子叶深绿不抗病:子叶浅绿抗病:子叶浅绿不抗病=1:1:1:1 |
| 实验三 | 子叶深绿抗病(♀)×子叶浅绿不抗病(♂) | 子叶深绿抗病:子叶深绿不抗病:子叶浅绿抗病:子叶浅绿不抗病=1:1:1:1 |
(1)上述诱变育种的原理是
(2)科研人员推测抗花叶病基因为
(3)根据表中实验
(4)实验一F1中的子叶浅绿抗病植株自交,F2成熟植株中子叶深绿抗病:子叶深绿不抗病:子叶浅绿抗病:子叶浅绿不抗病:子叶黄色抗病:子叶黄色不抗病为
(5)现有一株白花(dd)突变大豆,为研究D/d基因的位置,用该突变株做父本,与7-单体(仅缺失一条7号染色体,其余正常)紫花纯合子杂交得F1,统计F1的花色及比例。若F1中
(1)科学家获得了4号染色体的ps-2基因隐形突变体甲,表现为雄性不育,在杂交育种时,选育雄性不育植株的优点是
(2)科研人员用EMS诱变野生型番茄,获得雄性不育的突变体乙(甲、乙均只有一对基因与野生型不同)。下表为3个不同番茄杂交组合及其子代的表型及比例。请回答:
| 组合序号 | 亲本组合 | 后代的表型及比例 |
| 一 | 野生型×突变体甲 | 全为雄性可育(杂种1) |
| 二 | 野生型×突变体乙 | 全为雄性可育(杂种2) |
| 三 | 杂种1×杂种2 | 全为雄性可育 |
(3)已知4号染色体上的A基因可以指导植酸合成,不能合成植酸的会死亡。现有A基因缺失25个碱基对产生的A25-基因,效果未知。将基因型为AA25-的植物自交得到F1,提取F1各植株的DNA后进行PCR,选择的正向引物(从左到右)与A25-缺失的碱基配对,反向引物(从右到左)在其下游0.5kb处。将PCR产物进行电泳,发现各组产物电泳结果均具有条带,原因是
(4)用基因组编辑技术将一个A基因导入到基因型为A25-A25-的6号染色体上,A25-与导入的A基因之间的遗传
组别 | 亲本(P) | 子一代(F1) |
实验一 | 野生型和甲植株 | 高秆不抗病 |
实验二 | 野生型和乙植株 | 高秆不抗病 |
实验三 | 甲植株和乙植株 | 高秆不抗病:高秆抗病=1:1 |
回答下列问题:
(1)从实验
(2)从实验三可知,控制抗病与不抗病这对相对性状的基因
(3)实验一中的得到F1的基因型有
(4)要利用三组实验得到的F1来培育纯合的矮秆抗病水稻,最好选择
(1)对籼稻品种Z利用γ射线进行
(2)选取不同类型的籼稻进行杂交实验,结果见下表。
亲本 | F1表现型 | |
组合一 | Z×E-1 | 均与Z品种穗颈长度相似(短穗颈) |
组合二 | Z×E-2 | |
组合三 | E-1×E-2 |
②初步判断E-1和E-2所携带的长穗颈突变基因e1、e2为非等位基因,理由是
③el、e2分别控制穗颈不同节间的伸长,将上表中的
(3)已知粳稻的5号染色体上存在长穗颈突变基因e,科研人员欲将该基因转育到籼稻Z中,用于对e1、e2基因的进一步定位。
①将粳稻和籼稻Z先杂交再
②IR与E-2杂交的F1表现为短穗颈,与E-1杂交的F1表现为长穗颈。请在下图中标注出e1和e2基因,以说明el、e2与e的位置关系
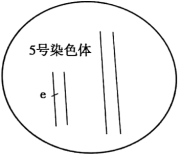
(4)若要利用E-1、E-2获得同时携带e1、e2两种突变基因且能稳定遗传的纯合品系,请结合(2)的相关信息尝试写出育种流程

回答下列问题:
(1)该植物配子中的所有染色体可称为
(2)通过实验一与实验二
(3)若让实验一F1相互杂交,与F1相比,子代中M基因频率
(4)实验三的亲本基因型为
(1)以Bt毒蛋白为有效成分的农药长期使用,会导致害虫种群
(2)以家蚕为材料,研究昆虫抗药性遗传特性。测定生命力大致相同的多品种家蚕幼虫致死50%的Bt浓度(LC50),结果如下:
家蚕品种 | A | B | C | D | E | F | G | H |
LC50(mg/L) | 59.4 | 37.5 | 94.2 | 118.6 | 118.6 | 118.7 | 129.0 | >375 |
1研究人员欲选择敏感型和抗药型家蚕为亲本进行杂交实验,据上表结果,最好选择家蚕品种是
②杂交实验及结果如下表:
亲本 | 实验一 ♀抗药型 ×♂敏感型 | 实验二 ♀敏感型 ×♂抗药型 | 实验三 实验一的 F1×抗药型 | 实验四 实验二的 F1×敏感型 | 实验五 实验一的 F1自交 | 实验六实验二的F1自交 | |
子代 | 抗药型 | 0 | 0 | 67 | 0 | 37 | 34 |
敏感型 | 80 | 80 | 83 | 150 | 113 | 116 | |
实验一和实验二的结果表明,家蚕抗药型是由位于
(3)Bt毒蛋白与家蚕幼虫的中肠细胞特异性受体结合,使细胞膜穿孔,细胞裂解,幼虫死亡。为从分子水平上阐明家蚕产生抗药性的原因,研究人员从实验一亲本

起始密码子 | 终止密码子 | 酪氨酸密码子 |
AUG/GUG | UAA/UAG | UAU/UAC |
从分子水平推测家蚕抗药性产生的原是敏感型受体基因中插入3个碱基对,导致
(4)依据Bt受体基因设计的靶向sgRNA,可以与限制酶Cas9结合,定点切除受体基因的39个核苷酸。sgRNA和Cas9mRNA的混合物注射到
(5)基因编辑技术能精确靶向修饰生物体基因组特定位点,人为改造生物体的遗传信息。请从利或弊某一方面说明该技术的应用是一把“双刃剑”:
表1
| 亲本组合 | 子代表型及比例 | |
| 一 | A×B | 一半存活且性状优良,一半生长一段时间后死亡 |
| 二 | A×C | 生长一段时间后全部死亡 |
| 亲本组合 | 总株数 | 成活株数 | 死亡株数 | |
| 一 | A×D | 20 | 20 | 0 |
| 二 | C×D | 20 | 20 | 0 |
| 三 | D×E | 19 | 19 | 0 |
| 四 | E×F | 20 | 11 | 9 |
(2)表1实验结果说明同时含有
(3)据表2分析,D的基因型为
(4)品种B的商品性状优良经济价值高,育种中科研人员先将B自交、分离出与D基因型一致的个体,再与A杂交,其后代
10 . I某昆虫的性别决定方式为XY型,用纯合猩红眼、驼胸雌性个体与纯合紫眼、野生胸雄性个体杂交,F1雌性个体和雄性个体全表现为野生眼、野生胸。F1雌雄个体杂交,F2的表型及个体数量(单位:只)如表所示。
性状 | 野生眼 野生胸 | 野生眼 驼胸 | 猩红眼 野生胸 | 猩红眼 驼胸 | 紫眼 野生胸 | 紫眼 驼胸 | 白眼 野生胸 | 白眼 驼胸 |
F2雌性个体 | 283 | 19 | 20 | 78 | 0 | 0 | 0 | 0 |
F2雄性个体 | 145 | 8 | 9 | 39 | 139 | 11 | 10 | 40 |
(1)昆虫眼色的遗传受2对独立遗传的等位基因的控制,其依据是
(2)控制野生胸与驼胸的基因为D/d,控制眼色的基因中与D/d位于同一对同源染色体上的基因是
II果蝇的体色由多对基因控制,野生型果蝇为灰体。现有黄体、黑体和黑檀体三种体色的果蝇单基因突变体(只有一对基因与野生型果蝇不同,不考虑互换和其他突变)。为探究果蝇体色基因的位置及显隐性关系,进行了下列实验。
亲本 | F1的表型 | |
实验一 | 黄体(雌)×野生型(雄) | 雌性均为灰体,雄性均为黄体 |
实验二 | 黑体(雌)×黑檀体(雄) | 均为灰体 |
实验三 | 黄体(雌)×黑檀体(雄) | 雌性均为灰体,雄性均为黄体 |
(4)根据实验三不能确定黑檀体基因的位置,理由是


