1 . 水稻叶色遗传可以归为CAPI体系:C为色素原基因,A为色素原活化基因,P为色素叶部表达基因,三基因同时表达叶色为紫色,否则为绿色,I为色素叶部表达基因的抑制基因,相对应隐性基因用c、a、p、i表示。将纯合紫叶品系甲与纯合绿叶品系乙、丙、丁、戊杂交得到F1,F1自交得到F2,实验结果见下表:
(1)控制叶色的4对等位基因的遗传____ (填“遵循”或“不遵循”)自由组合定律,判断依据是____ 。杂交组合④F1测交后代的表型及比例为____ 。
(2)粳稻和籼稻杂交子代具有高产等杂种优势,但通过传统去雄操作来进行人工杂交育种比较困难。科研人员利用γ射线诱变从正常可育系粳稻中获得了纯合反式光敏核雄性不育系,其育性受一对隐性核基因控制,表现为短日照败育,长日照可育。为大规模获得高产杂交水稻种子用于农业生产,可将____ 间行种植,在____ 日照条件下培育成植株并自然受粉,收获____ 植株的种子。
(3)研究发现该反式光敏核雄性不育系在短日照下仍有一定的自交结实率,导致培育的杂交种纯度低。为解决该问题,可选择纯合紫叶反式光敏核雄性不育系粳稻与纯合绿叶品系籼稻杂交,并在子代的秧苗期内剔除____ 色叶秧苗即可。该育种成功的前提条件是纯合绿叶品系籼稻具有____ 基因。
组合 | P | F1 | F2 |
① | 甲×乙 | 绿叶 | 绿叶∶紫叶 = 3∶1 |
② | 甲×丙 | 绿叶 | 绿叶∶紫叶 = 13∶3 |
③ | 甲×丁 | 绿叶 | 绿叶∶紫叶 = 55∶9 |
④ | 甲×戊 | 绿叶 | 绿叶∶紫叶 = 229∶27 |
(2)粳稻和籼稻杂交子代具有高产等杂种优势,但通过传统去雄操作来进行人工杂交育种比较困难。科研人员利用γ射线诱变从正常可育系粳稻中获得了纯合反式光敏核雄性不育系,其育性受一对隐性核基因控制,表现为短日照败育,长日照可育。为大规模获得高产杂交水稻种子用于农业生产,可将
(3)研究发现该反式光敏核雄性不育系在短日照下仍有一定的自交结实率,导致培育的杂交种纯度低。为解决该问题,可选择纯合紫叶反式光敏核雄性不育系粳稻与纯合绿叶品系籼稻杂交,并在子代的秧苗期内剔除
您最近一年使用:0次
2024-05-29更新
|
71次组卷
|
2卷引用:(教研室提供)2024届河北省邯郸市高三下学期5月保温试题生物试卷
2 . 家蚕(2n=56)是古代劳动人民最早从野蚕“驯化”而来,通常家蚕的体色为较浅的素色,但偶有黑色斑的突变体出现。
(1)家蚕与野蚕(2n=54)杂交子一代的染色体数为____________ 研究发现子一代依然可育,使用显微镜观察____________ 细胞,发现____________ 个二价体(两条染色体联会),一个三价体(三条染色体联会)。
(2)为研究家蚕体色遗传规律,科研人员进行了如下实验。
实验一:用黑色斑(黑蚕)与素色(素蚕)进行正反交实验,统计结果如下表所示。
根据以上杂交结果不能确定这对相对性状的显隐性,应该选择F1代_______________ 相互交配,若子代______________ ,则该性状表现为显性。
实验二:将F1代黑蚕相互交配及F2代黑蚕相互交配,统计结果如下表所示。
据此可知,黑色斑与素色这对相对性状中,_________ 为显性性状。推测存在____________ 才导致后代出现特殊的分离比。
(3)综合以上实验统计,F1→F3代黑蚕作母本时蚕受精卵的孵化率_______________ 。
(4)已知家蚕的黄血对白血是显性,控制该性状的基因位于2号染色体上(该基因不存在纯合致死现象),科研人员进行了如下实验;
亲本杂交→选择F1代个体杂交→统计F2代表现型及比例
(①/②) (③/④) (⑤/⑥)
①P∶黑蚕白血×素蚕黄血 ②P∶黑蚕黄血×素蚕白血
③F1∶黑蚕黄血×素蚕黄血 ④F1∶黑蚕黄血×素蚕白血
⑤F2∶黑蚕黄血:黑蚕白血:素蚕黄血=1∶1∶2 ⑥F2∶黑蚕黄血∶素蚕白血=1∶1
以上实验组合__________ 可说明黑色斑(素色)基因也在2号染色体上,判断依据是:____________ 。(只需从F2代的性状表现描述即可)
(1)家蚕与野蚕(2n=54)杂交子一代的染色体数为
(2)为研究家蚕体色遗传规律,科研人员进行了如下实验。
实验一:用黑色斑(黑蚕)与素色(素蚕)进行正反交实验,统计结果如下表所示。
交配方式 | 受精卵数 | F1代 | |
黑蚕 | 素蚕 | ||
(素♀×黑♂) | 730 | 363 | 364 |
(黑♀×素♂) | 737 | 323 | 317 |
根据以上杂交结果不能确定这对相对性状的显隐性,应该选择F1代
实验二:将F1代黑蚕相互交配及F2代黑蚕相互交配,统计结果如下表所示。
交配方式 | 受精卵数 | F2代/F3代 | |
黑蚕 | 素蚕 | ||
F1代黑蚕相互交配 | 715 | 253 | 125 |
F2代黑蚕相互交配 | 728 | 208 | 102 |
据此可知,黑色斑与素色这对相对性状中,
(3)综合以上实验统计,F1→F3代黑蚕作母本时蚕受精卵的孵化率
(4)已知家蚕的黄血对白血是显性,控制该性状的基因位于2号染色体上(该基因不存在纯合致死现象),科研人员进行了如下实验;
亲本杂交→选择F1代个体杂交→统计F2代表现型及比例
(①/②) (③/④) (⑤/⑥)
①P∶黑蚕白血×素蚕黄血 ②P∶黑蚕黄血×素蚕白血
③F1∶黑蚕黄血×素蚕黄血 ④F1∶黑蚕黄血×素蚕白血
⑤F2∶黑蚕黄血:黑蚕白血:素蚕黄血=1∶1∶2 ⑥F2∶黑蚕黄血∶素蚕白血=1∶1
以上实验组合
您最近一年使用:0次
名校
3 . 大豆花叶病毒会严重降低大豆的产量和品质。为预防抗病大豆品种甲(抗甲)的抗病能力减弱。科研人员用EMS诱变感病大豆,获得了新的抗病品种乙(抗乙)科研人员利用甲、乙品种进行杂交实验,结果如下表:
(1)基因突变具有的特点有______________________________ (答出3点)。
(2)据表中数据分析,品种甲的抗病性状属于_______________ 性状,品种乙的抗病性状属于_______________ 性状,且品种甲和品种乙都是______________________________ (纯合子/杂合子)。
(3)抗病品种乙是易感品种经诱变获得的。理论上抗病品种乙的抗性基因位置有3种可能情况:
I.品种乙的抗性基因与品种甲的抗性基因“分别位于两对同源染色体上”;
Ⅱ.品种乙的抗性基因与品种甲的抗性基因“位于同一对染色体上的不同位点上(不考虑交叉互换)”;
Ⅲ.品种乙的抗性基因与品种甲的抗性基因“位于同一对染色体上的相同位点上(在甲品种抗性基因相同的位点上发生突变产生另一等位基因)"。
将品种甲和品种乙杂交得F1,再自交得到F2。若为情况Ⅰ,则F2中抗病和易感个体的比例是_______________ ;若为情况Ⅱ,则F2中抗病和易感个体的比例是_______________ ;若为情况Ⅲ,则F2中抗病和易感个体的比例是___________________ 。
组别 | 亲本组合 | F1 | F2 | ||
抗病 | 易感病 | 抗病 | 易感 | ||
实验一 | 抗甲×易感 | 16 | 0 | 256 | 82 |
实验二 | 抗乙×易感 | 0 | 18 | 111 | 340 |
(1)基因突变具有的特点有
(2)据表中数据分析,品种甲的抗病性状属于
(3)抗病品种乙是易感品种经诱变获得的。理论上抗病品种乙的抗性基因位置有3种可能情况:
I.品种乙的抗性基因与品种甲的抗性基因“分别位于两对同源染色体上”;
Ⅱ.品种乙的抗性基因与品种甲的抗性基因“位于同一对染色体上的不同位点上(不考虑交叉互换)”;
Ⅲ.品种乙的抗性基因与品种甲的抗性基因“位于同一对染色体上的相同位点上(在甲品种抗性基因相同的位点上发生突变产生另一等位基因)"。
将品种甲和品种乙杂交得F1,再自交得到F2。若为情况Ⅰ,则F2中抗病和易感个体的比例是
您最近一年使用:0次
2022-07-15更新
|
254次组卷
|
2卷引用:四川省德阳市2021-2022学年高一下学期期末生物试题
4 . 被誉为“世界杂交水稻之父”的袁隆平,利用“野败”成功培育出了雄性不育水稻。杂交水稻主要是利用水稻雄性不育系(花粉不育)作为遗传工具,并形成了雄性不育系、恢复系和保持系三个核基因纯合品系的三系配套的第一代杂交水稻育种模式。
(1)杂交水稻普遍具有杂种优势,即杂种子一代在一种或多种性状上优于两个纯合亲本的现象。杂交水稻的培育工作中雄性不育品系的作用至关重要,因为其可免去大面积繁育制种时______ 操作,节省成本并保证杂交种子的纯度。
(2)雄性不育由细胞质不育基因S和核中隐性基因rf决定,即基因型(S)rfrf表现为雄性不育,而相应的细胞质基因N和细胞核中显性基因Rf都会使水稻恢复育性。三系配套杂交水稻有两个隔离的种植区(如下图),提示:杂交中细胞质基因不随花粉参与受精。
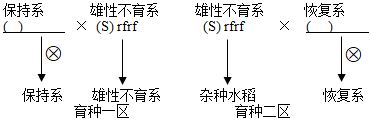
育种一区和二区除了种植雄性不育系,还分别间隔种植保持系和恢复系,通过自然杂交和自交,使三系得以保存,并在二区的雄性不育植株上收获了杂交水稻种子。写出保持系的基因型______ ,恢复系的基因型______ 。
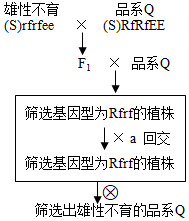
(3)某水稻品系Q是培育杂交水稻的优秀原材料,但表现为雄性可育,不便于直接用于制备杂交水稻。品系Q基因型为(S)RfRfEE,E基因与Rf基因位于非同源染色体上。雄性不育系的基因型为(S)rfrfee。
若要最大效率地培育出基因型为(S)rfrfEE的品系Q雄性不育系,可采用右图方法实现,框内为连续多代回交。图中回交亲本a是______ (雄性不育系/品系Q)。推算回交2次产生的F4中,基因型为EE的个体比例为______ 。最后所有F1植株自交产生(S)rfrfEE的比例是______ 。
(4)袁隆平团队培育出了光温敏雄性不育纯合新品种,即短日照平温条件下雄性可育,长日照高温条件下雄性不育,从而实现了两系配套的第二代杂交水稻。与恢复系混种,该品种植株上可收获全部是推广种植杂种的是______ (夏季/秋季)的成熟稻。这个实例也说明了植物的生长、发育和繁殖等生命活动的调节,除了激素调节外,还有______ 调节和______ 调控,它们是相互作用、协调配合的。
(1)杂交水稻普遍具有杂种优势,即杂种子一代在一种或多种性状上优于两个纯合亲本的现象。杂交水稻的培育工作中雄性不育品系的作用至关重要,因为其可免去大面积繁育制种时
(2)雄性不育由细胞质不育基因S和核中隐性基因rf决定,即基因型(S)rfrf表现为雄性不育,而相应的细胞质基因N和细胞核中显性基因Rf都会使水稻恢复育性。三系配套杂交水稻有两个隔离的种植区(如下图),提示:杂交中细胞质基因不随花粉参与受精。
育种一区和二区除了种植雄性不育系,还分别间隔种植保持系和恢复系,通过自然杂交和自交,使三系得以保存,并在二区的雄性不育植株上收获了杂交水稻种子。写出保持系的基因型
(3)某水稻品系Q是培育杂交水稻的优秀原材料,但表现为雄性可育,不便于直接用于制备杂交水稻。品系Q基因型为(S)RfRfEE,E基因与Rf基因位于非同源染色体上。雄性不育系的基因型为(S)rfrfee。
若要最大效率地培育出基因型为(S)rfrfEE的品系Q雄性不育系,可采用右图方法实现,框内为连续多代回交。图中回交亲本a是
(4)袁隆平团队培育出了光温敏雄性不育纯合新品种,即短日照平温条件下雄性可育,长日照高温条件下雄性不育,从而实现了两系配套的第二代杂交水稻。与恢复系混种,该品种植株上可收获全部是推广种植杂种的是
您最近一年使用:0次
2022-03-15更新
|
1918次组卷
|
4卷引用:辽宁省铁岭市六校2021-2022学年高三3月月考生物试题
5 . 小麦是雌雄同花植物。科研人员偶然发现一株高秆雄性不育小麦(甲),其雄性不育受显性基因M控制,另有一株矮秆可育小麦(乙),株高由A、a基因控制。科研人员进行了以下实验:
杂交一:甲×乙→F1矮秆雄性不育:矮秆雄性可育=1∶1
杂交二:F1中的矮秆雄性不育×高秆雄性可育(野生型)→F2矮秆雄性可育:高秆雄性不育=1:1回答下列问题:
(1)A、a基因的根本区别是_________ ,其显性现象的表现形式是_________ 。
(2)杂交一实验中甲作为_________ (父本/母本)。根据杂交一、二实验结果可知,甲和乙的基因型分别为_________ 、_________ ,两对等位基因的遗传_________ (遵循/不遵循)自由组合定律。
(3)假设让实验二的F2自由交配,后代中高秆雄性可育植株占_________ 。
(4)研究发现所有的雄性不育植株都特异性地携带了TRIM序列,且M及其等位基因的编码区相同;为研究该序列与雄性不育之间的关系,科研人员在野生型小麦中获得了M等位基因的启动子并利用Ti质粒构建表达载体,部分结构如下图所示,GFP表达后会发出绿色荧光。将载体转入小麦幼胚,获得转基因植株,对植株表型和M基因表达情况进行鉴定。
(1)表中①处转入的载体组成为_________ (从a~f中选择),②处植株表型为_________ 。
(2)在存活的转基因小麦花药中特异性检测到绿色荧光。根据实验结果推测导致雄性不育的原因是_________ 。
杂交一:甲×乙→F1矮秆雄性不育:矮秆雄性可育=1∶1
杂交二:F1中的矮秆雄性不育×高秆雄性可育(野生型)→F2矮秆雄性可育:高秆雄性不育=1:1回答下列问题:
(1)A、a基因的根本区别是
(2)杂交一实验中甲作为
(3)假设让实验二的F2自由交配,后代中高秆雄性可育植株占
(4)研究发现所有的雄性不育植株都特异性地携带了TRIM序列,且M及其等位基因的编码区相同;为研究该序列与雄性不育之间的关系,科研人员在野生型小麦中获得了M等位基因的启动子并利用Ti质粒构建表达载体,部分结构如下图所示,GFP表达后会发出绿色荧光。将载体转入小麦幼胚,获得转基因植株,对植株表型和M基因表达情况进行鉴定。

实验材料 | 转入载体组成I、II | 植株表型 | M基因表达情况 |
雄性不育小麦 | - | 雄性不育 | 仅在花药发育早期表达,在根、 茎、叶、雌蕊中均不表达 |
野生型小麦 | - | 可育 | 在花药发育各时期均不表达,在 根、茎、叶、雌蕊中均不表达 |
野生型小麦 | ad | 死亡 | - |
野生型小麦 | ①________ | ②________ | 仅在花药发育早期表达,在 根、茎、叶、雌蕊中均不表达 |
(2)在存活的转基因小麦花药中特异性检测到绿色荧光。根据实验结果推测导致雄性不育的原因是
您最近一年使用:0次
6 . 玉米为二倍体,玉米的卵细胞会进行有丝分裂,形成卵细胞和极核,精子与两个极核融合后发育形成胚乳,研究人员将基因型为Rr的玉米进行处理获得的突变体甲如图所示(数字代表染色体序号)。回答下列问题:

(1)正常基因型为Rr的玉米进行自交,获得的玉米籽粒胚乳的基因组成有________ 种,其基因组成为________________ 。突变体甲的形成是________________ 引起染色体结构变异(易位)。
(2)紫色RR(♂)×黄色rr(♀),F1胚乳为紫色,反交时F1胚乳为杂色。对胚乳颜色形成的机制有人作出了以下两种推测。
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。结果如下表。
表中基因组成为RRrr的胚乳产生的原因是________________ ,表中结果支持推测________ (填“一”“二”或“一和二”)。
(1)正常基因型为Rr的玉米进行自交,获得的玉米籽粒胚乳的基因组成有
(2)紫色RR(♂)×黄色rr(♀),F1胚乳为紫色,反交时F1胚乳为杂色。对胚乳颜色形成的机制有人作出了以下两种推测。
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。结果如下表。
杂交组合 | 部分F1胚乳 | ||
基因组成 | 颜色 | ||
I | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
II | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
您最近一年使用:0次
7 . 果蝇是常见的遗传学实验材料,请根据以下材料回答相关问题:


材料一:果蝇的红眼与白眼是一对相对性状,控制红眼(W+)与白眼(W)的基因位于X染色体上,红眼为显性性状;
材料二:用X射线诱变使果蝇X染色体上的W+基因移接到IV号染色体上,W+基因在IV号染色体上不能表达;
材料三:果蝇的眼是由多个小眼组成的复眼;
材料四:基因组成为XW+XW的果蝇经X射线照射后发育为红、白两色镶嵌的花斑眼果蝇。
(1)果蝇的体细胞中含有__________ 个染色体组,每个染色体组中含有一组__________ 染色体。
(2)材料二中提到的X射线诱导果蝇产生的变异类型是_______________ 。
(3)为什么基因型为XW+XW的果蝇眼睛颜色表现为花斑状?请根据上述材料提出合理假设:___________ 。
材料一:果蝇的红眼与白眼是一对相对性状,控制红眼(W+)与白眼(W)的基因位于X染色体上,红眼为显性性状;
材料二:用X射线诱变使果蝇X染色体上的W+基因移接到IV号染色体上,W+基因在IV号染色体上不能表达;
材料三:果蝇的眼是由多个小眼组成的复眼;
材料四:基因组成为XW+XW的果蝇经X射线照射后发育为红、白两色镶嵌的花斑眼果蝇。
(1)果蝇的体细胞中含有
(2)材料二中提到的X射线诱导果蝇产生的变异类型是
(3)为什么基因型为XW+XW的果蝇眼睛颜色表现为花斑状?请根据上述材料提出合理假设:
您最近一年使用:0次
8 . 回答下列(一)(二)小题
(一)假设a、B为玉米的优良基因,现有AABB、aabb两个品种,控制两对相对性状的基因位于两对同源染色体上,实验小组用不同方法进行了实验(见图),请回答下列问题

(1)过程①⑧育种方法是______ 育种,最大优点是能提高______ ,在短时间内获得更多的优良变异类型
(2)过程⑤使用的试剂是秋水仙素,它可作用于正在分裂的细胞抑制______ 的形成
(3)过程②③④育种方法运用的原理是______ ,基因型aaB_的类型经④后,子代中aaBB所占比例是______
(4)过程______ (填序号)应用了单倍体育种的方法,最大的优点是_________
(二)已知下述实验结果,请回答:颜色由两对基因(A和a, B和b)控制,其中一对基因控制色素的合成,另一对基因控制颜色的深浅,其花的颜色与基因型的对应关系见下表,请回答下列问题

(1)纯合白花植株和纯合红花植株做亲本进行杂交,产生的子一代花色全是红色,则亲代白花的基因型是______
(2)探究两对基因(A和a, B和b)的遗传是否符合基因的自由组合定律,某课题小组选用基因型为AaBb的植株进行测交。
实验步骤:第一步:对基因型为AaBb的植株进行测交。
第二步:观察并统计子代植株花的颜色和比例。
预期结果及结论
(2)①若子代花色及比例为______ ,则两对基因的遗传符合基因的自由组合定律,可表示为如图第一种类型(竖线表示染色体,黑点表示基因在染色体上的位置)。
②若子代植株花色出现其他分离比,则两对基因的遗传不符合基因的自由组合定律。请在表示方框中补充其它两种类型。

____________
(3)述两对基因的遗传符合自由组合定律,则基因型为AaBb的植株自交后代中,粉花植株的基因型有_______ 种。
(4)等位基因A和a影响花瓣的大小,基因型AA表现为大花瓣,Aa表现为小花瓣,aa表现为无花瓣,另有一对等位基因R和r影响花瓣的颜色,基因型RR和Rr表现为红色花瓣,rr表现为无色花瓣。两个植株双杂合子杂交,则下一代基因型_______ 种 ,表现型有_______ 种。
(一)假设a、B为玉米的优良基因,现有AABB、aabb两个品种,控制两对相对性状的基因位于两对同源染色体上,实验小组用不同方法进行了实验(见图),请回答下列问题
(1)过程①⑧育种方法是
(2)过程⑤使用的试剂是秋水仙素,它可作用于正在分裂的细胞抑制
(3)过程②③④育种方法运用的原理是
(4)过程
(二)已知下述实验结果,请回答:颜色由两对基因(A和a, B和b)控制,其中一对基因控制色素的合成,另一对基因控制颜色的深浅,其花的颜色与基因型的对应关系见下表,请回答下列问题
(1)纯合白花植株和纯合红花植株做亲本进行杂交,产生的子一代花色全是红色,则亲代白花的基因型是
(2)探究两对基因(A和a, B和b)的遗传是否符合基因的自由组合定律,某课题小组选用基因型为AaBb的植株进行测交。
实验步骤:第一步:对基因型为AaBb的植株进行测交。
第二步:观察并统计子代植株花的颜色和比例。
预期结果及结论
(2)①若子代花色及比例为
②若子代植株花色出现其他分离比,则两对基因的遗传不符合基因的自由组合定律。请在表示方框中补充其它两种类型。
(3)述两对基因的遗传符合自由组合定律,则基因型为AaBb的植株自交后代中,粉花植株的基因型有
(4)等位基因A和a影响花瓣的大小,基因型AA表现为大花瓣,Aa表现为小花瓣,aa表现为无花瓣,另有一对等位基因R和r影响花瓣的颜色,基因型RR和Rr表现为红色花瓣,rr表现为无色花瓣。两个植株双杂合子杂交,则下一代基因型
您最近一年使用:0次
解题方法
9 . (一)2019年,袁隆平被授予“共和国勋章”,以表彰他在杂交水稻研究领域作出的杰出贡献。请根据所学知识,回答有关某品种水稻(2N=24, 两性花,花多且小,自花授粉并结种子)育种的问题。
(二)科研人员将另外两种栽培水稻品系(甲和乙)进行下图所示杂交,培育抗赤霉菌感染的水稻品系。
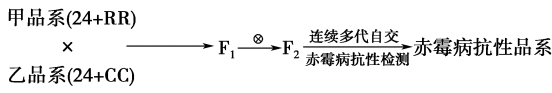
染色体组成为24+RR的水稻品系甲中,除含有水稻的24条染色体外,还具有两条来自一种野生稻的R染色体,R染色体携带抗赤霉病基因。染色体组成为24+CC的水稻品系乙中,除含有水稻的24条染色体外,另有两条来自另一种野生稻的C染色体。
(1)杂交育种的原理是_____ , 袁隆平找到的雄性不育的水稻,在进行杂交操作时的优势是______ 。
(2)水稻的抗稻瘟病性状(R/r)与抗倒伏性状(T/t)均由一对等位基因控制且独立遗传。现有抗稻瘟病易倒伏植株与易感稻瘟病抗倒伏植株杂交,F1中仅有抗稻瘟病易倒伏与易感稻瘟病易倒伏两类植株且比例为1∶1,则亲本基因组为________ ,技术人员在对基因型为rrTt的该品种水稻的幼苗用秋水仙素处理时,偶尔出现了一株基因型为RrrrTTtt的植株,该植株自交后代的性状分离比例是(只写出比例,不用写性状)______ 。
(3)该水稻花序的正常和异常是由一对等位基因控制的相对性状。某显性植株X自交,F1表现为正常花序∶异常花序=1∶1.取F1正常花序植株的花粉进行离体培养,获得的幼苗用秋水仙素处理后都是异常花序植株。由此推测________ 是显性性状,植株X自交的子代性状分离比为1∶1的原因可能是______ 。
(4)据图分析,F1的体细胞中染色体数为_______ , 染色体组成为_______ 。
(5)据图可知,甲、乙两品系之间并没有出现生殖隔离,因为它们的F1______ 。
(6)需从F2选择具有_____ 性状的个体,进行多代自交以及赤霉病抗性检测,最终获得赤霉病抗性品系,该品系具有赤霉病抗性是由于_____ 。
(二)科研人员将另外两种栽培水稻品系(甲和乙)进行下图所示杂交,培育抗赤霉菌感染的水稻品系。
染色体组成为24+RR的水稻品系甲中,除含有水稻的24条染色体外,还具有两条来自一种野生稻的R染色体,R染色体携带抗赤霉病基因。染色体组成为24+CC的水稻品系乙中,除含有水稻的24条染色体外,另有两条来自另一种野生稻的C染色体。
(1)杂交育种的原理是
(2)水稻的抗稻瘟病性状(R/r)与抗倒伏性状(T/t)均由一对等位基因控制且独立遗传。现有抗稻瘟病易倒伏植株与易感稻瘟病抗倒伏植株杂交,F1中仅有抗稻瘟病易倒伏与易感稻瘟病易倒伏两类植株且比例为1∶1,则亲本基因组为
(3)该水稻花序的正常和异常是由一对等位基因控制的相对性状。某显性植株X自交,F1表现为正常花序∶异常花序=1∶1.取F1正常花序植株的花粉进行离体培养,获得的幼苗用秋水仙素处理后都是异常花序植株。由此推测
(4)据图分析,F1的体细胞中染色体数为
(5)据图可知,甲、乙两品系之间并没有出现生殖隔离,因为它们的F1
(6)需从F2选择具有
您最近一年使用:0次
10 . 黄瓜是二倍体植物,茎、叶上有有毛和无毛之分,果实的表面有有无果刺、有无果瘤之分,相关杂交实验如表。
(1)杂交实验一中,控制茎、叶上有毛/无毛的基因位于__________ (填“核染色体”或“细胞质”)。某同学重复实验一,在亲本杂交时由于去雄不完全导致F2中有毛:无毛=7:2,推测杂交失误发生在__________ (填“正交”或“反交”)过程中。
(2)根据杂交实验二推测,控制__________ (填“有毛或有刺”“无毛或无刺”)性状的基因能抑制控制有瘤性状基因的表达。
(3)分析杂交实验二看出,有毛和有刺、无毛和无刺总是相伴而生。为此某同学提出了两种解释模型,请将模型加以完善。
模型一:“毛”和“刺”由一对等位基因控制,相同的基因型在茎、叶部位表现为“毛”,在果实表面表现为“刺”,是__________ 的结果。
模型二:“毛”和“刺”由两对等位基因控制,__________ ,且在F1产生配子过程中不发生交叉互换。
假定模型二成立,不考虑有瘤、无瘤性状及其基因,在重复杂交实验二时,若对发育成F1的一个受精卵进行某种诱变处理,若该处理引起的基因突变只突变成该基因的等位基因(不产生新基因),且该处理不影响配子及子代生活力,将该受精卵发育成的植株F1自交:若该处理诱发了F1毛和刺相关基因中的一个基因发生了基因突变,则F2有种__________ 表现型;若该处理诱发了F1受精卵中同源染色体上一对等位基因的互换(带有相关基因的相应染色体片段的断裂、移接和互换),则F2有__________ 种表现型。
| 实验 | 亲本 | F1表现型 | F2表现型及比例 |
| 杂交实验一 | 正交:♀有毛×♂无毛 反交:♂有毛×♀无毛 | 有毛 | 有毛:无毛=3:1 |
| 杂交实验二 | 正交:♀有毛、有瘤、有刺×♂无毛、无瘤、无刺 反交:♂有毛、有瘤、有刺×♀无毛、无瘤、无刺 | 有毛、有瘤、有刺 | 有毛、有瘤、有刺:有毛、无瘤、有刺:无毛、无瘤、无刺=9:3:4 |
(2)根据杂交实验二推测,控制
(3)分析杂交实验二看出,有毛和有刺、无毛和无刺总是相伴而生。为此某同学提出了两种解释模型,请将模型加以完善。
模型一:“毛”和“刺”由一对等位基因控制,相同的基因型在茎、叶部位表现为“毛”,在果实表面表现为“刺”,是
模型二:“毛”和“刺”由两对等位基因控制,
假定模型二成立,不考虑有瘤、无瘤性状及其基因,在重复杂交实验二时,若对发育成F1的一个受精卵进行某种诱变处理,若该处理引起的基因突变只突变成该基因的等位基因(不产生新基因),且该处理不影响配子及子代生活力,将该受精卵发育成的植株F1自交:若该处理诱发了F1毛和刺相关基因中的一个基因发生了基因突变,则F2有种
您最近一年使用:0次
2023-06-15更新
|
144次组卷
|
2卷引用:河南省开封市等2地2022-2023学年高三下学期五月期末考试理综生物试题


