名校
1 . 土壤盐碱化是我国最严重的生态环境问题之一,严重影响着农业生产。其中苏打盐碱地由于其含盐量高、高pH胁迫以及土壤通透性差,致使植物难以生长。水稻是我国主要的粮食作物之一,其生存能力强,盐碱地种植水稻是一项实现盐碱地资源高效利用的有利措施,可达到恢复生态与发展粮食经济的双重目的。据此,我国研究人员探究了苏打盐碱胁迫对水稻光合特性的影响,结果如下表,根据结果回答以下问题:
(1)水稻叶片叶绿素含量测定时,可先提取叶绿体色素,再进行测定。提取叶绿体色素时,选择___________ 作为提取液
(2)该实验的自变量为___________ ,气孔导度是否为该实验中净光合作用速率的限制因素?___________ (填“是”或“否”)。判断依据是___________ 苏打盐碱胁迫与叶绿素含量的关系是___________ 。
(3)脱落酸在植物应对环境胁迫中发挥重要的作用,为探究脱落酸(ABA)对水稻抗性的影响,科学家以D-4和J-51两种品系的水稻为实验材料,进行了一系列实验,结果如图。活性氧物质包括超氧自由基、H2O2等,活性氧的过量积累会对植物体造成多方面的损伤,能破坏植物体内的蛋白质、膜质、DNA、RNA等,同时植物体内的其他细胞组分也受到严重损伤,导致植物的多个生理代谢紊乱。_____________ .推测ABA的作用机理为____________ ,从而提高水稻的耐受性。
处理 | 叶绿 素a | 叶绿 素b | 叶绿 素a/b | 净光合作用速率 [μmol/(m2·s)] | 气孔导 度[μmol/(m2·s)] | 胞间CO2浓度 (μL/L) | 产量 (g/盆) |
对照组 | 1.05 | 0.18 | 5.78 | 22.08 | 1068.63 | 316.61 | 109.19 |
轻度胁迫 | 0.95 | 0.16 | 6.01 | 20.18 | 1045.28 | 332.40 | 93.98 |
中度胁迫 | 0.90 | 0.12 | 7.34 | 15.45 | 996.52 | 338.85 | 71.46 |
重度胁迫 | 0.82 | 0.08 | 10.42 | 10.81 | 952.12 | 343.44 | 53.48 |
(1)水稻叶片叶绿素含量测定时,可先提取叶绿体色素,再进行测定。提取叶绿体色素时,选择
(2)该实验的自变量为
(3)脱落酸在植物应对环境胁迫中发挥重要的作用,为探究脱落酸(ABA)对水稻抗性的影响,科学家以D-4和J-51两种品系的水稻为实验材料,进行了一系列实验,结果如图。活性氧物质包括超氧自由基、H2O2等,活性氧的过量积累会对植物体造成多方面的损伤,能破坏植物体内的蛋白质、膜质、DNA、RNA等,同时植物体内的其他细胞组分也受到严重损伤,导致植物的多个生理代谢紊乱。

您最近一年使用:0次
7日内更新
|
36次组卷
|
2卷引用:2024届重庆市涪陵五中高三春第一次适应性考试生物试题
名校
2 . 农杆菌转化法可将外源基因导入到羊角蜜细胞中改变其遗传特性,转化操作前对野生农杆菌的Ti质粒进行改造(图甲所示)。转化成功的羊角蜜细胞进行植物组织培养时,利用不同浓度6-BA诱导不定芽形成,结果如图乙所示。下列分析不正确的是( )

| A.为了使外源基因在羊角蜜细胞中表达,则重组质粒中的启动子最好来源于羊角蜜细胞 |
| B.标记基因1可用于筛选成功导入T-DNA的植物细胞 |
| C.标记基因2与报告基因的转录模板链不在同一条DNA单链上 |
| D.图乙结果不能表明低浓度6-BA促进不定芽形成,高浓度6-BA抑制不定芽形成 |
您最近一年使用:0次
2024-05-29更新
|
100次组卷
|
2卷引用:2024届重庆市部分学校高三下学期5月巴渝名校联合考试生物试卷
名校
解题方法
3 . 某科研小组为研究细胞分裂素(CTK)对叶片面积的影响。将油菜豆幼苗去除根和幼芽制成插条后插入蒸馏水中,并用人工合成的细胞分裂素类似物PBA对插条进行不同处理,方法及结果如下表,下列有关叙述错误的是( )
| 组别 | 处理方法 | A叶面积相对值 | |
1 | 实验组 | PBA处理B | 1.7 |
| 对照组 | 等量蒸馏水处理B | 2.5 | |
2 | 实验组 | PBA处理A | 3.6 |
3 | 对照组 | 等量蒸馏水处理A | ? |
| 实验组 | PBA处理A,摘除B | 3.2 | |
| 对照组 | 等量蒸馏水处理A,摘除B | 2.3 | |
| A.制备插条时除去根和幼芽是为了防止内源CTK的干扰 |
| B.实验的自变量就是PBA处理的不同部位,组别2对照组的结果应该也在2.5左右 |
| C.组别1实验组的结果可能是PBA引起营养物质向处理部位运输所致 |
| D.根据结果推测,若摘除叶片A,叶片B的面积可能会减小 |
您最近一年使用:0次
4 . 独脚金内酯是一类新发现的植物激素。为了探究独脚金内酶的作用,以及与生长素在调控植物顶端优势中的相互关系,科研人员以拟南芥为实验材料,进行了相关实验。
(1)植物顶芽产生的生长素,通过__________ 运输到侧芽处,使顶芽生长素浓度降低从而促进其生长,侧芽处生长素浓度过高从而抑制其生长,这种现象称为顶端优势。
(2)研究发现独脚金内酯合成受阻或不敏感突变体均无顶端优势,但生长素水平正常。科研人员将上述两种突变体与野生型(W)进行嫁接试验,培养后植株形态如下表所示。
分析上表,第1组嫁接植株表现出顶端优势,推测独脚金内酯合成部位最可能在_________ ,其作用为影响侧枝的生长;第2组嫁接结果无顶端优势,可能的原因是突变体2为___________ (“独脚金内酯合成受阻突变体”或“独脚金内酯不敏感突变体”);第4组的嫁接结果最可能为_________ 。
(3)为进一步研究独脚金内酯与生长素的关系,科研人员作出如下假设:生长素沿主茎运输时,独脚金内酯会抑制侧芽的生长素向主茎运输,造成侧芽生长素浓度升高,从而形成顶端优势。下图为实验设计装置。________ (A或B)中添加适量生长素类似物处理主茎,在侧枝处施加一定量具有放射性的________ (“生长素类似物”或“独脚金内酯类似物”),一段时间后,检测主茎下端放射性强度。
②实验组:用生长素类似物对主茎进行与对照组相同处理,在侧枝处施加等量的与对照组相同的物质,在主茎另一端的固体培养基中施加适量的__________ ,一段时间后,检测主茎下端放射性强度。若实验结果为__________ ,则证明假设成立。
(1)植物顶芽产生的生长素,通过
(2)研究发现独脚金内酯合成受阻或不敏感突变体均无顶端优势,但生长素水平正常。科研人员将上述两种突变体与野生型(W)进行嫁接试验,培养后植株形态如下表所示。
组别 | 嫁接处理 | 结果(顶端优势) |
1 | 突变体1的地上部分+W的根 | 有 |
2 | 突变体2的地上部分+W的根 | 无 |
3 | 突变体2地上部分+突变体1的根 | 无 |
4 | 突变体1地上部分+突变体2的根 | ? |
(3)为进一步研究独脚金内酯与生长素的关系,科研人员作出如下假设:生长素沿主茎运输时,独脚金内酯会抑制侧芽的生长素向主茎运输,造成侧芽生长素浓度升高,从而形成顶端优势。下图为实验设计装置。

②实验组:用生长素类似物对主茎进行与对照组相同处理,在侧枝处施加等量的与对照组相同的物质,在主茎另一端的固体培养基中施加适量的
您最近一年使用:0次
名校
解题方法
5 . 植物生命活动调节中激素调节占据着非常重要的地位。水稻种子形成时,胚乳的发育和营养物质积累决定水稻的产量和品质。研究发现,胚乳形成过程中不同激素间的相互作用如图所示。下列分析错误的是( )

| A.多种植物激素协同调控籽粒长度 |
| B.油菜素内酯是能起调节作用的微量有机物 |
| C.生长素通过增大细胞体积促进籽粒长度增加 |
| D.油菜素内酯和细胞分裂素能促进籽粒变短变窄 |
您最近一年使用:0次
名校
解题方法
6 . 为研究桃树矮化的机理,研究人员对矮化品种F和正常株高品种Q分别施加赤霉素(GA3)后,测定枝条生长情况,并检测与生长相关蛋白Della的含量,结果如图所示。有资料显示,GA3与受体蛋白GID1结合后,GID1与Della形成复合物,进而通过特定途径影响Della的含量。下列分析不正确的是( )


| A.内源性GA3在植物的幼芽、幼根中均有合成 | B.相较于品种Q,品种F对GA3不敏感 |
| C.蛋白Della具有促进枝条生长的作用 | D.可通过降低GID1基因的表达得到矮化品种 |
您最近一年使用:0次
2024-05-06更新
|
135次组卷
|
3卷引用:2024届重庆市巴蜀中学校高三下学期高考适应性月考卷(九)生物试题
名校
解题方法
7 . GA(赤霉素)可上调PIN1(生长素转运蛋白)的表达水平,进而改变生长素的分布。某同学设计下表实验验证上述结论,实验材料、试剂有野生型拟南芥、PIN1缺陷型拟南芥(不能合成PIN1)、GA溶液、清水等。下列叙述错误 的是( )
实验组别 | 实验材料 | 实验处理 | 实验结果 |
对照组 | ① | ③ | 与对照组相比,实验组1中PIN1的表达水平上升,生长素分布在更靠近韧皮部一侧;对照组、实验组2的生长素分布情况接近 |
实验组1 | 野生型拟南芥 | GA溶液 | |
实验组2 | ② | ④ |
| A.表中①②分别为野生型拟南芥、PIN1缺陷型拟南芥 |
| B.表中③④分别为GA溶液、清水 |
| C.该实验的自变量为有无GA处理和植株的种类 |
| D.该实验的因变量为PIN1的表达水平、生长素的分布情况 |
您最近一年使用:0次
2024-05-06更新
|
378次组卷
|
4卷引用:2024届重庆市部分学校高三考前最后一卷生物试题
解题方法
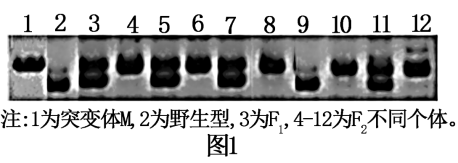
8 . 玉米是雌雄同株异花植物,研究人员利用纯合突变体M探究玉米花“性别决定”的分子机制。该突变体表现为雄穗中雌蕊发育,雄蕊退化,即雄穗雌性化。将M与野生型杂交,所得的F1全部表现为雄穗正常,F1个体间随机传粉,所得F2中雄穗正常:雄穗雌性化=108:35。请回答下列问题:
(1)野生型玉米雄穗中的雌蕊发育不久即退化消失,这一生命现象属于细胞的_____ 。
(2)玉米1号染色体上有一标记序列S,不同品种S序列长度不同。科研人员利用S序列对突变体M的雄穗雌性化基因进行定位,PCR扩增上述杂交实验的亲本、F1及F2若干个体的S序列,电泳结果如图1。_____ ,且F2中的_____ (填图中序号)均表现为雄穗正常。
(3)进一步研究发现,雄穗雌性化基因的定位区间中包含性别决定基因T(表达产物是抑制雌蕊发育的信号),推测M的表型是T基因突变所致。为验证这一推测,科研人员PCR扩增突变体M的T基因序列与野生型对比,该基因的非模板链部分序列及部分氨基酸的密码子如图2。_____ 。
②发育中的雌蕊会产生赤霉素,过量的赤霉素抑制雄蕊发育。由此可知,T基因突变使得玉米的内源赤霉素含量_____ ,最终表现为雄穗雌性化。
(4)科研人员向野生型玉米的T基因中插入片段使其功能完全丧失,构建了纯合突变体M1。突变体M1雌性化完全,不能产生有功能的花粉;但突变体M雌性化不完全,能产生少量有功能的花粉。为验证M是M1的等位突变体,请设计杂交实验并预期结果_____ 。
(1)野生型玉米雄穗中的雌蕊发育不久即退化消失,这一生命现象属于细胞的
(2)玉米1号染色体上有一标记序列S,不同品种S序列长度不同。科研人员利用S序列对突变体M的雄穗雌性化基因进行定位,PCR扩增上述杂交实验的亲本、F1及F2若干个体的S序列,电泳结果如图1。
(3)进一步研究发现,雄穗雌性化基因的定位区间中包含性别决定基因T(表达产物是抑制雌蕊发育的信号),推测M的表型是T基因突变所致。为验证这一推测,科研人员PCR扩增突变体M的T基因序列与野生型对比,该基因的非模板链部分序列及部分氨基酸的密码子如图2。

②发育中的雌蕊会产生赤霉素,过量的赤霉素抑制雄蕊发育。由此可知,T基因突变使得玉米的内源赤霉素含量
(4)科研人员向野生型玉米的T基因中插入片段使其功能完全丧失,构建了纯合突变体M1。突变体M1雌性化完全,不能产生有功能的花粉;但突变体M雌性化不完全,能产生少量有功能的花粉。为验证M是M1的等位突变体,请设计杂交实验并预期结果
您最近一年使用:0次
解题方法
9 . 渍害是因洪、涝积水或地下水位过度升高,导致作物根系长期缺氧进而对植株造成伤害。发生渍害时,油菜地上部分以有氧呼吸为主。地下部分细胞利用丙酮酸进行乙醇发酵,此代谢过程中需要乙醇脱氢酶的催化,促进氢接受体(NAD+)再生,从而使葡萄糖分解得以顺利进行。回答下列问题:
(1)渍害条件下乙醇脱氢酶活性越高的品种越_____ (耐渍害/不耐渍害)。
(2)以不同渍害能力的油菜品种为材料,经不同时长的渍害处理,测定相关生理指标并进行相关性分析,结果见下表。
注:表中数值为相关系数r,代表两个指标之间相关的密切程度。当|r|接近1时,相关越密切,越接近0时相关越不密切。
据表分析,与叶绿素含量呈负相关的指标是_____ 。已知渍害条件下光合速率显著下降,则蒸腾速率呈_____ 趋势。综合分析表内各指标的相关性,光合速率下降主要由_____ (气孔限制因素/非气孔限制因素)导致的,理由是_____ 。
(3)植物通过形成系列适应机制响应渍害。受渍害时,植物体内_____ (激素)大量积累,诱导气孔关闭,调整相关反应,防止有毒物质积累,提高植物对渍害的耐受力:渍害发生后,有些植物根系细胞通过_____ ,将自身某些薄壁组织转化腔隙,形成通气组织,促进氧气运输到根部,缓解渍害。
(1)渍害条件下乙醇脱氢酶活性越高的品种越
(2)以不同渍害能力的油菜品种为材料,经不同时长的渍害处理,测定相关生理指标并进行相关性分析,结果见下表。
光合速率 | 蒸腾速率 | 气孔导度 | 胞间CO2浓度 | 叶绿素含量 | |
光合速率 | 1 | ||||
蒸腾速率 | 0.95 | 1 | |||
气孔导度 | 0.99 | 0.94 | 1 | ||
胞间CO2浓度 | -0.99 | -0.98 | -0.99 | 1 | |
叶绿素含量 | 0.86 | 0.90 | 0.90 | -0.93 | 1 |
据表分析,与叶绿素含量呈负相关的指标是
(3)植物通过形成系列适应机制响应渍害。受渍害时,植物体内
您最近一年使用:0次
名校
10 . 涝胁迫会严重危害作物生长发育甚至导致作物死亡。为缓解涝胁迫,植物通过形成不定根和通气组织、节间快速伸长等方式维持生长。植物激素在调控上述生理和形态变化中发挥了重要作用。回答下列问题:
(1)涝胁迫诱导形成不定根的植物激素主要是__________ ,在细胞水平上其作用主要是___________________ (答两点),在黄瓜中,涝胁迫下积累的蔗糖诱导 PIN 以及 LBD基因表达,在消耗细胞内能量的前提下促进该激素的运输,诱导不定根的发生和形成。如果 PIN、LBD基因失去功能,与正常植株相比,在涝胁迫下黄瓜植株可能的变化是_______________________ 。
(2)发生涝胁迫后,水稻节间快速增长,为确定节间快速增长与细胞的伸长生长直接相关,可采用的鉴定方法是___________ 。赤霉素除具有促进细胞伸长外,主要作用还包括_________________________ (答2点)。
(3)已知乙烯是涝胁迫下诱导通气组织形成的重要植物激素,为验证外源或内源乙烯均会诱导玉米根系通气组织的形成,选择生长状况相同的玉米植株若干进行实验,请写出实验思路:____________________ 。
(1)涝胁迫诱导形成不定根的植物激素主要是
(2)发生涝胁迫后,水稻节间快速增长,为确定节间快速增长与细胞的伸长生长直接相关,可采用的鉴定方法是
(3)已知乙烯是涝胁迫下诱导通气组织形成的重要植物激素,为验证外源或内源乙烯均会诱导玉米根系通气组织的形成,选择生长状况相同的玉米植株若干进行实验,请写出实验思路:
您最近一年使用:0次


