1 . 在日本著名ACG作品《fate》系列中,剧中人物的发色的家族遗传方式往往令人浮想联翩,下面介绍其中一种假说:假设在fate平行宇宙中,人类的发色由三对互不连锁的等位基因决定,第一对基因为复等位基因,由B(决定黑色发色),G(决定墨绿色发色),a(决定白色发色组成);第二对基因决定橘红色发色(R-r);第三对基因(I-i)编码一种抑制蛋白,且对第一对基因的基因产物有显性抑制作用。黑色发色对橘红色和墨绿色发色有掩盖作用,当其体内只存在橘红色和墨绿色色素时,两者将发生反应生成紫色复合物,具体通路如下。阅读以下材料,回答下列小题。
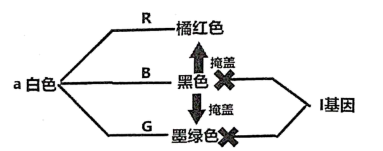
1.如图,远坂时臣(发色黑色)与远坂葵(发色墨绿色)育有两女,远坂凛发色黑色并在远坂家修习宝石魔法长大成人,而远坂樱初为黑色头发,但在7岁时过继到间桐家并改名为间桐樱,由于刻印虫的影响发色和瞳色逐渐变为紫色。另一方面,爱丽丝菲尔•冯•爱因兹贝伦(发色白色)与卫宫切嗣(发色黑色)缔结婚姻,并生育一女取名为伊利亚斯菲尔•冯•爱因兹贝伦,发色亦为白色。后卫宫切嗣夺取圣杯失败,收养一孤儿,取名卫宫士郎(发色橘红色)即为男主。已知远坂葵和爱丽丝菲尔冯爱因兹贝伦由于出自名门,其三对基因都为纯合,且卫宫切嗣的纳除黑色外不存在其他色素基因。下列关于个人基因型的相关分析中,正确的是?( )

2.承上题,又知爱丽丝菲尔•冯•爱因兹贝伦和卫宫切嗣第一对基因分别为BB和Ba,男主在第五次圣杯战争中与上题中的远坂凛、间桐樱和伊利亚斯菲尔•冯•爱因兹贝伦发生了一些“奇妙”的羁绊,下列关于卫宫士郎对于她们的选择中,错误的选项是?【不考虑基因突变,染色体变异等小概率事件】( )
1.如图,远坂时臣(发色黑色)与远坂葵(发色墨绿色)育有两女,远坂凛发色黑色并在远坂家修习宝石魔法长大成人,而远坂樱初为黑色头发,但在7岁时过继到间桐家并改名为间桐樱,由于刻印虫的影响发色和瞳色逐渐变为紫色。另一方面,爱丽丝菲尔•冯•爱因兹贝伦(发色白色)与卫宫切嗣(发色黑色)缔结婚姻,并生育一女取名为伊利亚斯菲尔•冯•爱因兹贝伦,发色亦为白色。后卫宫切嗣夺取圣杯失败,收养一孤儿,取名卫宫士郎(发色橘红色)即为男主。已知远坂葵和爱丽丝菲尔冯爱因兹贝伦由于出自名门,其三对基因都为纯合,且卫宫切嗣的纳除黑色外不存在其他色素基因。下列关于个人基因型的相关分析中,正确的是?( )
| A.可以推断远坂葵的基因型为rriiGG |
| B.刻印虫可能导致间桐樱体内B等位基因所在的染色体异染色质化或产生了组蛋白修饰改变,从而引起其发色的变化 |
| C.可以确定远坂凛的基因型有两对杂合,一对纯合 |
| D.可以推测伊利亚斯菲尔•冯•爱因兹贝伦和其母亲的白发性状的形成与第一对基因无关 |
| E.伊利亚在I-i基因上一定为杂合,在R-r基因上可能为杂合。 |
| A.可知卫宫士郎的基因型为Rriiaa或RRiiaa |
| B.如果卫宫士郎选择和间桐樱在一起,则他们的子女的发又可能有橘红色,紫色,白色的情况 |
| C.如果卫宫士郎选择和伊利亚在一起,且后代为白发的概率是1/2,则可推测后代发色的性状比为白色:黑色:橘红色=4:3:1 |
| D.无论卫宫士郎选择与哪位女主在一起,他都不能保证后代一定是有色发色(即非白色)。 |
您最近一年使用:0次
2 . 学习以下材料,回答(1)-(4)题。
动物细胞的非经典蛋白分泌途径
蛋白分泌是细胞间信息交流的重要途径。通常所指的蛋白分泌是经典分泌,即具有信号肽序列的分泌蛋白被信号肽识别因子识别后进入内质网,通过内质网-高尔基体运输释放,大多数分泌蛋白通过此途径分泌。研究发现,一些不含信号肽的蛋白可不依赖于经典分泌途径而被释放到细胞外,这些分泌途径统称为非经典蛋白分泌(UPS)。
UPS分为膜泡运输和非膜泡运输两大类。膜泡运输介导的UPS存在一个关键问题:缺乏信号肽的蛋白是如何进入膜泡中的?在内质网和高尔基体之间存在一种管泡状结构,称为内质网-高尔基体中间体(ERGIC)。在经典分泌途径中,ERGIC会对蛋白运输的方向进行选择:若蛋白是错误分选运输至ERCIC,其会产生反向运输的膜泡将蛋白运回内质网;对于正确分选的蛋白,其通过膜泡顺向运输至高尔基体。研究者发现了定位于ERGIC膜上的TMEDI0蛋白,缺乏信号肽的分泌蛋白通过结合细胞质中的HSP90A来帮助其发生去折叠,进而该蛋白与TMEDI0相互作用,诱导TMED10寡聚化形成蛋白通道。在HSP90B1的帮助下,TMED10蛋白与缺乏信号肽的分泌蛋白中一段由14个氨基酸组成的序列结合,促进该蛋白进入到ERGIC腔内,如图。最后该蛋白包裹进ERCIC膜形成的膜泡中被直接运送到细胞膜或进入分泌型自噬体,分泌型自噬体又可以直接和细胞膜融合或与分泌型溶酶体融合,最终将蛋白释放到细胞外。
(1)内质网、ERGIC、膜泡等多种细胞结构都有膜,这些膜共同构成细胞的__________ 。
(2)研究某种蛋白在细胞中分泌途径的方法有__________。
(3)有人认为ERGIC是细胞蛋白分泌过程中膜泡转运和导向的枢纽,依据是__________ 。
(4)根据文中信息,推测UPS存在的意义是__________ 。
动物细胞的非经典蛋白分泌途径
蛋白分泌是细胞间信息交流的重要途径。通常所指的蛋白分泌是经典分泌,即具有信号肽序列的分泌蛋白被信号肽识别因子识别后进入内质网,通过内质网-高尔基体运输释放,大多数分泌蛋白通过此途径分泌。研究发现,一些不含信号肽的蛋白可不依赖于经典分泌途径而被释放到细胞外,这些分泌途径统称为非经典蛋白分泌(UPS)。
UPS分为膜泡运输和非膜泡运输两大类。膜泡运输介导的UPS存在一个关键问题:缺乏信号肽的蛋白是如何进入膜泡中的?在内质网和高尔基体之间存在一种管泡状结构,称为内质网-高尔基体中间体(ERGIC)。在经典分泌途径中,ERGIC会对蛋白运输的方向进行选择:若蛋白是错误分选运输至ERCIC,其会产生反向运输的膜泡将蛋白运回内质网;对于正确分选的蛋白,其通过膜泡顺向运输至高尔基体。研究者发现了定位于ERGIC膜上的TMEDI0蛋白,缺乏信号肽的分泌蛋白通过结合细胞质中的HSP90A来帮助其发生去折叠,进而该蛋白与TMEDI0相互作用,诱导TMED10寡聚化形成蛋白通道。在HSP90B1的帮助下,TMED10蛋白与缺乏信号肽的分泌蛋白中一段由14个氨基酸组成的序列结合,促进该蛋白进入到ERGIC腔内,如图。最后该蛋白包裹进ERCIC膜形成的膜泡中被直接运送到细胞膜或进入分泌型自噬体,分泌型自噬体又可以直接和细胞膜融合或与分泌型溶酶体融合,最终将蛋白释放到细胞外。

(1)内质网、ERGIC、膜泡等多种细胞结构都有膜,这些膜共同构成细胞的
(2)研究某种蛋白在细胞中分泌途径的方法有__________。
| A.用放射性同位素标记氨基酸,追踪细胞中放射性物质出现的部位 |
| B.用荧光染料标记ERGIC膜蛋白,观察细胞中荧光的迁移路径 |
| C.用药物阻断内质网与高尔基体间的膜泡运输,检测蛋白在细胞内的分布 |
| D.构建TMED10基因缺失的突变细胞系,检测蛋白在突变细胞内的分布 |
(4)根据文中信息,推测UPS存在的意义是
您最近一年使用:0次
2023-05-08更新
|
1110次组卷
|
2卷引用:2023届北京市朝阳区高三二模生物试题
名校
解题方法
3 . 阅读资料,回答下列小题:
叶绿体与线粒体间信号交流调控植物程序性细胞死亡
程序性细胞死亡(PCD)是生物体受遗传调控的自主细胞死亡现象,在植物生长发育和抵抗环境胁迫中起重要作用。植物在遭受各种生物或非生物胁迫时,体内ROS增加,ROS作为信号分子会增加线粒体膜的通透性,诱导细胞发生程序性死亡。
中国科学院遗传与发育生物学研究所李家洋研究组筛选出1个拟南芥细胞死亡突变体mod1,该突变体中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关。通过modl突变体克隆鉴定出MODI基因,该基因编码叶绿体中的脂肪酸合酶,负调控植物PCD。
近年来的研究表明,线粒体在PCD中起核心作用。一个有趣的问题是,叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD?为探明其中的作用机制,他们针对modl突变体构建了其T-DNA插入突变体库,从中筛选出能够抑制modl细胞死亡和ROS积累表型的抑制突变体,并克隆了这些抑制突变体对应的抑制基因,该基因与线粒体电子传递链复合体I(mETC复合体I)的组装及活性有关。modl是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体I的功能缺失所恢复,由此暗示植物细胞中存在叶绿体与线粒体之间的信号交流调控PCD。
最近,研究组通过大规模筛选modl突变体的抑制突变体,克隆了3个新的抑制基因pINAD-MDH、DiT l和mMDH 1。此3个基因分别编码质体定位的NAD+依赖的苹果酸脱氢酶、叶绿体被膜定位的二羧酸转运蛋白1和线粒体定位的苹果酸脱氢酶1,突变后都可抑制mod1中ROS的积累及PCD的发生。通过对这些基因进行深入的功能分析,他们论证了苹果酸从叶绿体到线粒体的转运,对线粒体中ROS的产生及随后PCD的诱导起重要作用(如图)。
1.根据上述图文信息,下列关于植物细胞中叶绿体与线粒体之间的信号交流调控PCD机制的叙述,错误的是( )
2.根据文中信息,下列证据中能够支持上述机制的是( )
叶绿体与线粒体间信号交流调控植物程序性细胞死亡
程序性细胞死亡(PCD)是生物体受遗传调控的自主细胞死亡现象,在植物生长发育和抵抗环境胁迫中起重要作用。植物在遭受各种生物或非生物胁迫时,体内ROS增加,ROS作为信号分子会增加线粒体膜的通透性,诱导细胞发生程序性死亡。
中国科学院遗传与发育生物学研究所李家洋研究组筛选出1个拟南芥细胞死亡突变体mod1,该突变体中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关。通过modl突变体克隆鉴定出MODI基因,该基因编码叶绿体中的脂肪酸合酶,负调控植物PCD。
近年来的研究表明,线粒体在PCD中起核心作用。一个有趣的问题是,叶绿体中导致ROS产生的信号是否可传递到线粒体中激发ROS产生并最终诱导PCD?为探明其中的作用机制,他们针对modl突变体构建了其T-DNA插入突变体库,从中筛选出能够抑制modl细胞死亡和ROS积累表型的抑制突变体,并克隆了这些抑制突变体对应的抑制基因,该基因与线粒体电子传递链复合体I(mETC复合体I)的组装及活性有关。modl是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体I的功能缺失所恢复,由此暗示植物细胞中存在叶绿体与线粒体之间的信号交流调控PCD。
最近,研究组通过大规模筛选modl突变体的抑制突变体,克隆了3个新的抑制基因pINAD-MDH、DiT l和mMDH 1。此3个基因分别编码质体定位的NAD+依赖的苹果酸脱氢酶、叶绿体被膜定位的二羧酸转运蛋白1和线粒体定位的苹果酸脱氢酶1,突变后都可抑制mod1中ROS的积累及PCD的发生。通过对这些基因进行深入的功能分析,他们论证了苹果酸从叶绿体到线粒体的转运,对线粒体中ROS的产生及随后PCD的诱导起重要作用(如图)。
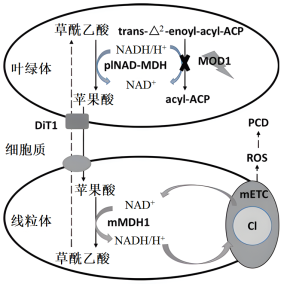
1.根据上述图文信息,下列关于植物细胞中叶绿体与线粒体之间的信号交流调控PCD机制的叙述,错误的是( )
| A.叶绿体中MOD1功能缺失导致NADH/H+在叶绿体中大量积累,草酰乙酸在pINAD-MDH作用下被氧化为苹果酸 |
| B.苹果酸通过DiT1转运到细胞质中,并进一步转移到线粒体 |
| C.在线粒体中,mMDH1将苹果酸转化为草酰乙酸,同时NADH/H+水平升高 |
| D.mETC复合体I活性增加,产生过量ROS,引发PCD |
| A.拟南芥细胞死亡突变体mod1中存在明显的ROS积累,暗示ROS的过量积累与该突变体的细胞死亡表型相关 |
| B.mod1是叶绿体中脂肪酸合酶的突变体,其PCD表型却能被线粒体中mETC复合体1所恢复 |
| C.抑制突变体的三个抑制基因突变后均可抑制MOD1发生PCD |
| D.苹果酸也可以由线粒体运输到叶绿体 |
您最近一年使用:0次
2024-06-01更新
|
72次组卷
|
2卷引用:天津市南开中学2023—2024学年高三下学期5月模拟生物试题
解题方法
4 . 果蝇的红眼和白眼是一对相对性状,其中红眼是显性性状,由位于 X染色体上的 W 基因控制。选用若干红眼雌果蝇和白眼雄果蝇作为亲代杂交,每代果蝇再随机交配,统计各代雌、雄果蝇中 w基因频率的变化,曲线如下图所示。若每代能繁殖出足够数量的个体,下列叙述错误的是( )
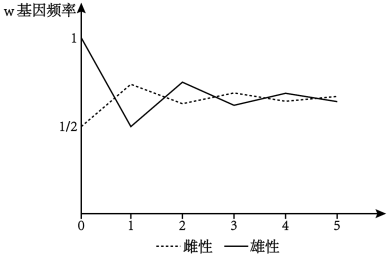
| A.亲本果蝇的基因型为 XWXw和 XwY |
| B.每一代果蝇群体中w 基因频率均为 2/3 |
| C.每代雌性与雄性中 W 基因频率之和的1/2与下一代雌性个体中 W的基因频率相等 |
| D.子三代雌性中 w 基因频率为 11/16,雄性中 w基因的频率为5/8 |
您最近一年使用:0次
名校
解题方法
5 . 科研人员欲利用pET22b质粒将L-天冬酰胺酶基因(如图1)导入宿主菌(对氨苄青霉素敏感)中,以构建高效表达L-天冬酰胺酶的菌株。该质粒上含有氨苄青霉素抗性基因(AmpR)和LacZ基因及一些酶切位点,其结构如图2。LacZ基因编码的产生的半乳糖苷酶可以分解X-gal产生蓝色物质,使菌落周围呈蓝色,否则菌落为白色。利用感受态法将基因表达载体导入宿主菌后(转化成功率并不是100%),可将转化后的宿主菌接种在含氨苄青霉素和X-gal的固体培养基上,以此筛选出成功导入重组质粒的宿主菌。下列叙述错误 的是( )
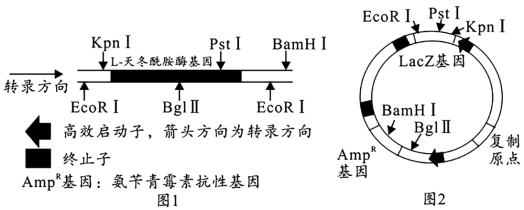
| A.用BglⅡ和BamHⅠ对目的基因和质粒进行切割,成功导入重组质粒的宿主菌在含氨苄青霉素的固体培养基上不能生长 |
| B.可用EcoRⅠ对目的基因和质粒进行切割,周围呈白色的菌落为成功导入重组质粒的宿主菌,均可产生L-天冬酰胺酶 |
| C.可用EcoRⅠ和KpnⅠ对目的基因和质粒进行切割,其酶切产物不能直接混合用于基因表达载体的构建 |
| D.可用EcoRⅠ和KpnⅠ对目的基因和质粒进行切割,周围呈蓝色的菌落为成功导入重组质粒的宿主菌 |
您最近一年使用:0次
解题方法
6 . 人类某遗传病受一对基因T、t控制,人类ABO血型受3个复等位基因ⅠA、ⅠB、i控制,位于另一对染色体上。A血型的基因型有ⅠAⅠA、ⅠAi,B血型的基因型有ⅠBⅠB、ⅠBi,AB血型的基因型为ⅠAⅠB,O血型的基因型为ii。两个家系成员的性状表现如图,Ⅱ-3和Ⅱ-5均为AB血型,Ⅱ-4和Ⅱ-6均为O血型。下列说法错误的是( )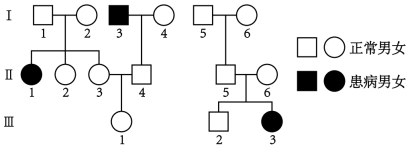
| A.该遗传病是由位于常染色体上的隐性致病基因控制的 |
| B.Ⅱ-3的基因型为TtⅠAⅠB的概率为2/3,Ⅱ-4的基因型一定为Ttii |
| C.如果Ⅲ-1与Ⅲ-2婚配,为预防生出患该遗传病的子代,最好进行遗传咨询 |
D.若Ⅲ-1与Ⅲ-2生育一个正常女孩,该女孩为O血型且携带致病基因的概率为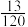 |
您最近一年使用:0次
7 . 果蝇受精卵中性染色体组成及发育情况如下表所示,研究果蝇的红白眼性状(由位于X染色体上的等位基因 Aa 控制)遗传时发现,一只白眼雌果蝇与红眼雄果蝇杂交,F1中红眼雌果蝇:白眼雌果蝇:红眼雄果蝇:白眼雄果蝇 =4:1:1:4,F1自由交配得到F2,不考虑基因突变和染色体互换,下列说法错误的是( )
受精卵中性染色体组成 | 发育情况 |
XX、XXY | 雌性,可育 |
XY、XYY | 雄性。可育 |
XXX、YO(无X染色体)、YY | 胚胎期致死 |
XO(无Y染色体) | 雄性,不育 |
| A.亲本白眼雌果蝇最多能产生4种类型的配子 |
| B.F1中红眼雌果蝇的基因型为XAXa、XAXaY |
| C.F1中雌果蝇产生Xa配子的概率为1/3 |
| D.F2中雌果蝇的基因型最多有5种可能 |
您最近一年使用:0次
名校
解题方法
8 . 某卵原细胞正常染色体和倒位染色体由于同源区段的配对及交换,在减数分裂过程中形成倒位环,产生一条含双着丝粒桥的染色体,如图所示,“×”表示交换位点。无着丝粒片段会因水解而丢失,双着丝粒桥在两着丝粒之间随机发生断裂。不考虑其他突变和基因被破坏的情况,下列说法错误的是( )
| A.次级卵母细胞中一条染色单体上的基因不可能是ebcAB |
| B.卵细胞的基因组成最多有10种可能 |
| C.若卵细胞的基因组成是Dcbe,则第二极体可能是ABCDE、adcbe和E |
| D.若卵细胞基因组成是EDc,则双着丝粒桥断裂位点一定发生在c、d之间 |
您最近一年使用:0次
9 . 根据突变表型,将上述3种不同的节律突变基因分别命名为“无节律”、“长周期”和“短周期”。利用杂交获得在两条X染色体上分别携带有不同突变的雌蝇,并对其羽化节律进行测试,结果如表所示。阅读材料完成下列小题:
1.互补实验常用于确定两个突变是否影响同一个基因。以下可以说明两个隐性突变并不位于同一基因的是( )
2.请根据表中所示的数据,下列结论错误的是( )
| 基因型组成 | |||
| 第一条X染色体 | 第二条X染色体 | 测试次数 | 节律周期时长统计(hr) |
| 野生型 | 野生型 | 4 | 24.4±0.5 |
| 无节律 | 无节律 | 4 | 无节律 |
| 短周期 | 短周期 | 5 | 19.5±0.4 |
| 长周期 | 长周期 | 4 | 28.6±0.5 |
| 无节律 | 野生型 | 8 | 25.2±0.4 |
| 短周期 | 野生型 | 5 | 21.9±0.4 |
| 长周期 | 野生型 | 5 | 25.6±0.5 |
| 短周期 | 无节律 | 6 | 19.5±0.4 |
| 长周期 | 无节律 | 8 | 30.6±1.3 |
| 短周期 | 长周期 | 6 | 22.9±0.4 |
| A.杂交后代表现出正常表型 |
| B.杂交后代表现出突变特征 |
| C.杂交后代表现出中间表型 |
| D.杂交后代表型更严重 |
| A.无节律突变相对于野生型等位基因,表现为隐性突变 |
| B.即使存在野生型等位基因,短周期突变仍可影响节律 |
| C.无节律突变对短周期突变和长周期突变而言均为显性 |
| D.短周期突变对长周期突变完全显性 |
您最近一年使用:0次
名校
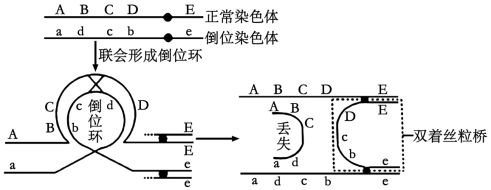
10 . 家蝇Y染色体由于某种影响断成两段,含s基因的小片段移接到常染色体获得XY'个体,不含s基因的大片段丢失。含s基因的家蝇发育为雄性,只含一条X染色体的雌蝇胚胎致死,其他均可存活且繁殖力相同。M、m是控制家蝇体色的基因,灰色基因M对黑色基因m为完全显性。如图所示的两亲本杂交获得F1,从F1开始逐代随机交配获得Fn。不考虑互换和其他突变,下列说法错误的是( )
| A.F1至Fn中所有个体均可由体色判断性别 |
| B.F1代中黑身∶灰身=1∶2 |
| C.F2雄性个体中,含有2条X染色体的个体占3/4 |
| D.F1至Fn代,雌性个体所占比例逐代降低 |
您最近一年使用:0次


