名校
解题方法
1 . 葡萄被誉为世界四大水果之首,它营养丰富、用途广泛、色美、气香、味可口,是果中佳品。光照条件下,叶肉细胞中O2与CO2竞争性结合五碳糖,O2与五碳糖结合后经一系列反应释放 CO2的过程称为光呼吸。回答下列问题:
(1)________ 循环将CO2还原成糖。据材料可推测光呼吸过程中 O2与五碳糖结合在叶肉细胞的________ 中进行。葡萄属于高光呼吸型的植物,为降低光呼吸对葡萄光合产物的消耗,研究者向葡萄叶面喷施不同浓度的光呼吸抑制剂SoBS溶液,相应的光合作用强度和光呼吸强度见下表。
注:表中光合作用强度即总光合速率,SoBS溶液处理对叶片呼吸作用的影响忽略不计。据表分析,该实验的光合作用强度用________ 来表示。喷施浓度为300mg/L SoBS 溶液对葡萄增产________ (填“有利”或“不利”)。为探究SoBS 溶液利于增产的最适喷施浓度,应在________ mg/L之间再设置多个浓度梯度进一步进行实验。
(2)葡萄叶光合产物主要以________ 形式提供给果实。
(3)大型封闭式气调冷藏库(充入氮气替换部分空气),延长了葡萄保鲜时间、增加了果农收益。气调冷藏库中葡萄的需氧呼吸受到抑制,原因是________ 。该库中葡萄细胞的糖酵解________ (填“会”或“不会”)受到抑制。气调冷藏库配备的过滤装置及时清除葡萄细胞产生的________ (激素),可延长葡萄保鲜时间。
(1)
SoBS 浓度(mg/L) | 0 | 100 | 200 | 300 | 400 | 500 | 600 |
光合作用强度(CO2μmol•m-2•s-1) | 18.9 | 20.9 | 20.7 | 18.7 | 17.6 | 16.5 | 15.7 |
光呼吸强度(CO2μmol•m-2•s-1) | 6.4 | 6.2 | 5.8 | 5.5 | 5.2 | 4.8 | 4.3 |
(2)葡萄叶光合产物主要以
(3)大型封闭式气调冷藏库(充入氮气替换部分空气),延长了葡萄保鲜时间、增加了果农收益。气调冷藏库中葡萄的需氧呼吸受到抑制,原因是
您最近一年使用:0次
2024-04-25更新
|
282次组卷
|
4卷引用:2024届浙江省稽阳联谊二模生物试题
名校
2 . 某实验小组将小麦种子横切两部分:“含胚半粒”和“无胚半粒”,用清水浸泡后,分别把两部分及等量缓冲液加入4支试管中,并按下表中的操作进行实验。保温一段时间后,取各试管中等量的上清液,加碘检测,结果见下表。请据表分析下列说法错误的是( )
注:“-”为不添加相应物质,“+”为浅蓝色,“+++”为深蓝色。
试管编号 | 半粒种子(10粒) | 赤霉素(10-6g/mL) | 赤霉素抑制剂(5×10-4mol/L) | 检测结果(蓝色深浅) |
1 | 有胚 | - | - | + |
2 | 无胚 | - | - | +++ |
3 | 无胚 | 0.2mL | - | + |
4 | 无胚 | 0.2mL | 0.2mL | +++ |
| A.1、2组结果可以说明小麦种子产生淀粉酶与胚有关 |
| B.3组与2、4组结果不同,说明赤霉素可以诱导无胚部分产生淀粉酶 |
| C.1、3组检测结果相同可能因为有胚部分的赤霉素诱导淀粉酶的合成 |
| D.根据表格可推测,小麦种子在成熟过程中赤霉素的含量逐渐增多 |
您最近一年使用:0次
2023-02-11更新
|
54次组卷
|
2卷引用:河南省鹤壁市高中2022-2023学年高二下学期第一次段考生物试题
名校
3 . 矮化栽培是现代苹果生产的趋势,为研究矮化苹果(M9)砧木致矮机理,研究者进行了系列实验。
(1)生长素在植物茎尖从形态学上端向下端的运输称为_________ 运输;细胞分裂素主要在植物体的根尖合成并运至地上部分,通过促进细胞分裂进而促进植物生长。
(2)研究者将富士苹果(非矮化)、M9和八棱海棠(非矮化)三种植物进行嫁接,如图1。一段时间后,检测富士-M9-八棱海棠不同部位PIN基因(IAA运输载体基因)的表达情况,结果如图2。
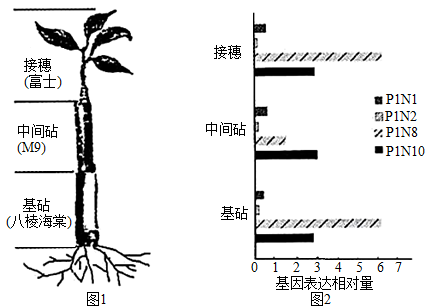
实验结果显示,在富士-M9-八棱海棠中,表达量较高的基因有两种,其中__________ 可能与M9致矮密切相关。
(3)向富士-M9-八棱海棠的接穗叶片或基砧根部施加NAA(生长素类调节剂,其运输载体与IAA相同)后,以未做处理的富士-M9-八棱海棠作为对照,检测相关指标如下表。
注:“+”代表检测指标的数值,“+”越多,数值越大。
①补充表中“?”处的实验步骤:____________________ ;
②与对照组相比,接穗叶片施加NAA组的基砧中(NAA+IAA)总含量几乎无差异,推测中间砧__________ (填“促进”或“抑制”)了生长素的运输。为证实这一推测,研究者将中间砧(M9)换成八棱海棠后,基砧中(NAA+IAA)总含量显著增加,这一结果__________ (支持/不支持)上述推测。与对照组相比,第3组的基砧中__________ 含量显著增加,推测基砧根部较高浓度的NAA促进了基砧根部该激素的合成。
③两实验组中,接穗叶片施加NAA组的接穗新梢平均长度较短,请你结合(2)(3)实验结果解释原因,以揭示M9砧木致矮机理:___________ 。
(1)生长素在植物茎尖从形态学上端向下端的运输称为
(2)研究者将富士苹果(非矮化)、M9和八棱海棠(非矮化)三种植物进行嫁接,如图1。一段时间后,检测富士-M9-八棱海棠不同部位PIN基因(IAA运输载体基因)的表达情况,结果如图2。
实验结果显示,在富士-M9-八棱海棠中,表达量较高的基因有两种,其中
(3)向富士-M9-八棱海棠的接穗叶片或基砧根部施加NAA(生长素类调节剂,其运输载体与IAA相同)后,以未做处理的富士-M9-八棱海棠作为对照,检测相关指标如下表。
| 组别 | 接穗新梢 平均长度 | 细胞分裂素含量 | (NAA+IAA)总含量 | ||
| 接穗叶片 | 基砧根部 | 接穗 | 基砧 | ||
| 1对照组 | + | + | ++ | ++ | + |
| 2接穗叶片施加NAA | + | + | ++ | +++ | + |
| 3? | +++ | +++ | ++++ | ||
①补充表中“?”处的实验步骤:
②与对照组相比,接穗叶片施加NAA组的基砧中(NAA+IAA)总含量几乎无差异,推测中间砧
③两实验组中,接穗叶片施加NAA组的接穗新梢平均长度较短,请你结合(2)(3)实验结果解释原因,以揭示M9砧木致矮机理:
您最近一年使用:0次
2022-07-04更新
|
262次组卷
|
4卷引用:河北省五个一名校联盟2022-2023学年高三上学期摸底考试生物试题
4 . 植物的不同部位激素种类和含量存在一定的差异(详情见下表所示)。植物的生长发育是由多种激素相互协调、共同调节的。请回答下列问题:
(1)目前公认的植物激素共有五大类,即生长素、赤霉素、细胞分裂素、脱落酸和乙烯,这些激素中在生物体内分布最广的是___________ ,理由是___________ 。
(2)根据表中信息,生长素和赤霉素二者的共同效应主要是通过促进细胞的伸长,从而促进植物的生长,而不是促进细胞数目的增多,理由是___________ 。
(3)进一步的研究得知,细胞分裂素可通过抑制叶绿素、核酸和蛋白质的降解,从而抑制叶片衰老。为证明细胞分裂素有延缓叶片衰老的作用,现设计以下实验。请完善相关步骤,并分析实验结果。
实验原理:因叶绿素的逐渐降解而导致叶片失绿变黄是叶片衰老的最显著特点,因此,可通过用细胞分裂素来涂抹新鲜的离体叶片,记录叶片失绿变黄所需的时间来证明。
实验步骤:
第一步:选取同种植物、大小和发育状况相同的叶片随机均分成甲、乙两组。
第二步:在甲组叶片上涂抹一定浓度的细胞分裂素,在乙组叶片的___________ 。
第三步:观察记录甲、乙两组叶片失绿变黄所需时间。
实验的结果及结论:___________ ,细胞分裂素有延缓叶片衰老的作用。
| 部位 | 激素的相对浓度 | |||
| 生长素 | 赤霉素 | 细胞分裂素 | 脱落酸 | |
| 茎尖 | +++ | +++ | +++ | - |
| 幼叶 | +++ | +++ | - | - |
| 伸长茎 | ++ | ++ | - | - |
| 侧芽 | + | + | - | - |
| 成熟叶 | + | + | - | +++ |
| 注:“+++”表示含量高;“++”表示含量中等;“+”表示含量低;“-”表示无。 | ||||
(2)根据表中信息,生长素和赤霉素二者的共同效应主要是通过促进细胞的伸长,从而促进植物的生长,而不是促进细胞数目的增多,理由是
(3)进一步的研究得知,细胞分裂素可通过抑制叶绿素、核酸和蛋白质的降解,从而抑制叶片衰老。为证明细胞分裂素有延缓叶片衰老的作用,现设计以下实验。请完善相关步骤,并分析实验结果。
实验原理:因叶绿素的逐渐降解而导致叶片失绿变黄是叶片衰老的最显著特点,因此,可通过用细胞分裂素来涂抹新鲜的离体叶片,记录叶片失绿变黄所需的时间来证明。
实验步骤:
第一步:选取同种植物、大小和发育状况相同的叶片随机均分成甲、乙两组。
第二步:在甲组叶片上涂抹一定浓度的细胞分裂素,在乙组叶片的
第三步:观察记录甲、乙两组叶片失绿变黄所需时间。
实验的结果及结论:
您最近一年使用:0次
解题方法
5 . 为了研究光照和赤霉素(GA)对菟丝子幼苗缠绕寄主的影响,某兴趣小组用培养基培养菟丝子及苜蓿(菟丝子的寄主),分别向培养基中添加10-2μmol/L、10-3μmol/L、10-4μmol/L、10-5μmol/L、10-6μmol/L的赤霉素合成抑制剂,并给与适宜光照条件。回答下列问题:
(1)植物体内GA的合成部位主要是未成熟的种子、幼根和幼芽,其主要作用是_______ (答出1点即可)。
(2)实验结果表明,在加入10-2μmol/L、10-4μmol/L赤霉素合成抑制剂的培养基中,菟丝子分别表现出“没有发生缠绕”、“不缠绕到缠绕的过渡状态”。试推测,在加入10-6μmol/L赤霉素合成抑制剂的培养基中,菟丝子的缠绕情况表现为_______ (填“缠绕”、“不缠绕”或“不缠绕到缠绕的过渡状态”),原因是______________ 。
(3)某同学认为,该实验不能说明菟丝子缠绕的发生受光照影响,其理由是______________ 。
(4)有研究表明,光照会影响赤霉素合成及其代谢有关基因的表达,进而影响赤霉素含量。综上所述可知,植物生命活动的调节是______________ 共同作用的结果。
(1)植物体内GA的合成部位主要是未成熟的种子、幼根和幼芽,其主要作用是
(2)实验结果表明,在加入10-2μmol/L、10-4μmol/L赤霉素合成抑制剂的培养基中,菟丝子分别表现出“没有发生缠绕”、“不缠绕到缠绕的过渡状态”。试推测,在加入10-6μmol/L赤霉素合成抑制剂的培养基中,菟丝子的缠绕情况表现为
(3)某同学认为,该实验不能说明菟丝子缠绕的发生受光照影响,其理由是
(4)有研究表明,光照会影响赤霉素合成及其代谢有关基因的表达,进而影响赤霉素含量。综上所述可知,植物生命活动的调节是
您最近一年使用:0次
名校
6 . 生长素(IAA)是重要的植物生长调节剂。生长素运输渠道化理论认为:顶芽产生的生长素通过主茎运输,当主茎中生长素运输流饱和时,会限制侧芽合成的生长素外流,侧芽处于休眠状态,形成顶端优势。另有研究发现,细胞分裂素(CK)和独脚金内酯也参与侧芽生长调控,三者关系如图8所示,其中→表示促进,表示抑制,连续箭头表示运输路径。
(2)据图,在调控侧芽生长方面,生长素与细胞分裂素表现为_________ (协同/拮抗)作用,生长素与独脚金内酯表现为_________ (协同/拮抗)作用。
(3)对顶端优势进行解释时,生长素运输渠道化理论与图8所示机理的差别主要在于后者_____。
(4)人工打顶后,侧芽能快速生长甚至开花结实。生产中常采用打顶后及时涂抹生长素的方法,使叶片增多、叶面积增大,以提高烟草等植物叶片的产量。其原理是该方法可以_____ 。(编号选填)
①控制开花和结果的总量②减少叶片中有机物的输出③增加开花和结果的机会④增加叶片中有机物的积累
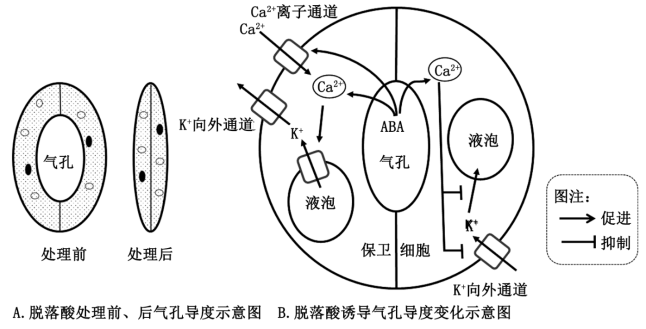
研究人员发现,干旱胁迫下,植物体通过分泌脱落酸(ABA)调节保卫细胞的生理变化,帮助植物抵抗逆境伤害。调节机制如图中A和B所示。
(6)根据图并结合已有知识,分析说明脱落酸(ABA)通过调节气孔导度帮助植物抵抗干旱胁迫的机制。______________ 。
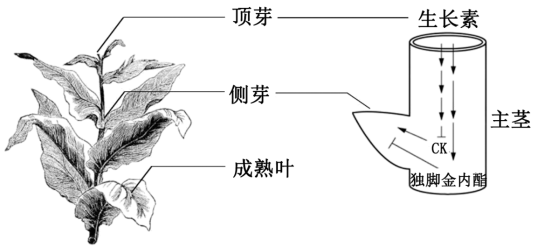
| A.侧芽处生长素合成加强 | B.侧芽处生长素外流加强 |
| C.侧芽处生长素合成减弱 | D.侧芽处生长素外流减弱 |
(2)据图,在调控侧芽生长方面,生长素与细胞分裂素表现为
(3)对顶端优势进行解释时,生长素运输渠道化理论与图8所示机理的差别主要在于后者_____。
| A.关注了激素运输对分布带来的影响 |
| B.关注了单一种类激素对植物的影响 |
| C.关注了多种激素对植物生命活动的调节 |
| D.关注了外界环境对植物生命活动的调节 |
(4)人工打顶后,侧芽能快速生长甚至开花结实。生产中常采用打顶后及时涂抹生长素的方法,使叶片增多、叶面积增大,以提高烟草等植物叶片的产量。其原理是该方法可以
①控制开花和结果的总量②减少叶片中有机物的输出③增加开花和结果的机会④增加叶片中有机物的积累
研究人员发现,干旱胁迫下,植物体通过分泌脱落酸(ABA)调节保卫细胞的生理变化,帮助植物抵抗逆境伤害。调节机制如图中A和B所示。
| A.赤霉素 | B.乙烯 | C.生长素 | D.细胞分裂素 |
(6)根据图并结合已有知识,分析说明脱落酸(ABA)通过调节气孔导度帮助植物抵抗干旱胁迫的机制。
您最近一年使用:0次
7 . 学习以下材料, 回答(1) ~(4)题。
铝对植物的毒害及植物的抗铝机制
铝是地壳中含量最丰富的金属元素, 地球上多达50%的可耕地为酸性土壤, 酸性条件下地壳中的铝以可溶性三价离子的形式被释放出来, 抑制植物根的生长发育。植物也通过一些机制减轻铝的毒害作用。
同时, 很多植物在进化过程中还形成了多种抗铝机制。小麦、拟南芥、大豆等植物根尖细胞存在苹果酸转运蛋白(ALMT), 铝离子可引发ALMT空间结构变化, 使其孔道打开, 细胞向外分泌苹果酸等有机酸可螯合根际土壤中的铝离子。再有, 铝毒害还可引起ALMT基因的表达量上升或转运蛋白在根中的重新分布。
有关植物对铝毒害的信号感知与调控机制的研究不断深入, 这些为未来开展作物分子育种设计和可持续农业发展提供了理论支撑。
(1)生长素_________ 运输, 称为极性运输。
(2)研究显示乙烯位于生长素调控上游, 下列支持该论点的证据有 。
(3)据文中信息, 分别阐释铝毒害对双子叶、单子叶植物根生长抑制的作用机制。
①双子叶植物(如拟南芥):_________ , 导致T区中生长素浓度较高, 根生长受抑制。
②单子叶植物(如玉米):_________ , 从而造成根生长受抑制。
(4)结合文中信息, 选择单子叶或双子叶作物之一, 提出培育耐铝作物的思路_______________ 。
铝对植物的毒害及植物的抗铝机制
铝是地壳中含量最丰富的金属元素, 地球上多达50%的可耕地为酸性土壤, 酸性条件下地壳中的铝以可溶性三价离子的形式被释放出来, 抑制植物根的生长发育。植物也通过一些机制减轻铝的毒害作用。
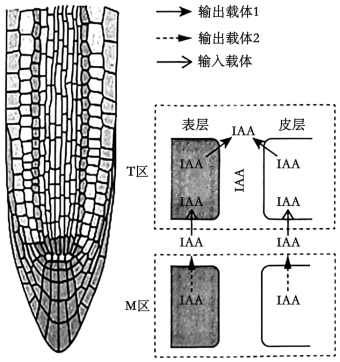
同时, 很多植物在进化过程中还形成了多种抗铝机制。小麦、拟南芥、大豆等植物根尖细胞存在苹果酸转运蛋白(ALMT), 铝离子可引发ALMT空间结构变化, 使其孔道打开, 细胞向外分泌苹果酸等有机酸可螯合根际土壤中的铝离子。再有, 铝毒害还可引起ALMT基因的表达量上升或转运蛋白在根中的重新分布。
有关植物对铝毒害的信号感知与调控机制的研究不断深入, 这些为未来开展作物分子育种设计和可持续农业发展提供了理论支撑。
(1)生长素
(2)研究显示乙烯位于生长素调控上游, 下列支持该论点的证据有 。
| A.乙烯处理后, 生长素输出载体2和输入载体的表达增加 |
| B.外源施加生长素极性运输阻断剂使植株呈明显的耐铝表型 |
| C.加入乙烯合成抑制剂, 可减弱铝毒害下T区生长素合成相关基因的表达 |
| D.铝毒害时, 乙烯受体突变体T区的生长素合成基因表达量低于野生型 |
(3)据文中信息, 分别阐释铝毒害对双子叶、单子叶植物根生长抑制的作用机制。
①双子叶植物(如拟南芥):
②单子叶植物(如玉米):
(4)结合文中信息, 选择单子叶或双子叶作物之一, 提出培育耐铝作物的思路
您最近一年使用:0次
名校
8 . 光合作用机理是作物高产的重要理论基础,光饱和点是指光合速率不再随光照强度增加时的光照强度,光补偿点是指光合过程中吸收的CO₂与呼吸过程中释放的( CO₂量相等时的光照强度。研究发现水稻野生型(WT)的产量和突变体(ygl)在不同栽培条件下产量有差异。
(1)测得两种水稻分别在弱光照和强光照条件下净光合速率的变化如下图1、图2所示:_________ (填“高”或“低”)有关。所有生物的生存,都离不开细胞呼吸,细胞呼吸的意义是_____________________ (答出2点即可)。
②据图分析,为了提高产量,在常年阳光充足、光照强度大的地区,更适合种植_____________ 水稻,依据是______________________ 。
③某研究者欲用希尔反应来测定两种水稻首蓿叶绿体的活力。希尔反应基本过程是将_________ (填“黑暗”或“光照”)中制备的离体叶绿体加到含有DCIP(氧化型)、蔗糖和 pH7.3磷酸缓冲液的溶液中并照光。水在光照下被分解,溶液中的DCIP 被还原,颜色由蓝色变成无色。氧化型 DCIP 在希尔反应中的作用,相当于____________ 在光反应中的作用。
(2)通常情况下,叶绿素含量与植物的光合速率成正相关。但上述研究表明,在强光照条件下,突变体(ygl) 水稻光合速率反而明显高于野生型(WT)。为进一步探究其原因,研究者在相同光照强度的强光条件下,测定了两种水稻的相关生理指标见下表:
注: RuBP 羧化酶是指催化( CO₂固定的酶; Vmax表示 RuBP 羧化酶催化的最大速率
①植物激素在植物光合作用中发挥重要作用,其中_________ (填激素名称)能够促进叶绿素的合成。植物生长发育的调控,是由________ 调控、激素调节和___________ 因素调节共同完成的。
②据表分析,在强光照条件下,突变体水稻光合速率反而明显高于野生型的原因是______________ 。
(1)测得两种水稻分别在弱光照和强光照条件下净光合速率的变化如下图1、图2所示:

②据图分析,为了提高产量,在常年阳光充足、光照强度大的地区,更适合种植
③某研究者欲用希尔反应来测定两种水稻首蓿叶绿体的活力。希尔反应基本过程是将
(2)通常情况下,叶绿素含量与植物的光合速率成正相关。但上述研究表明,在强光照条件下,突变体(ygl) 水稻光合速率反而明显高于野生型(WT)。为进一步探究其原因,研究者在相同光照强度的强光条件下,测定了两种水稻的相关生理指标见下表:
水稻材料 | 叶绿素(mg/g) | 类胡萝卜素 ( mg/g) | RuHP 羧化酶含量(单位: 略) | Vmax(单位: 略) |
WT | 4.08 | 0.63 | 4.6 | 129.5 |
ygl | 1.73 | 0.47 | 7.5 | 164.5 |
①植物激素在植物光合作用中发挥重要作用,其中
②据表分析,在强光照条件下,突变体水稻光合速率反而明显高于野生型的原因是
您最近一年使用:0次
2024-06-07更新
|
54次组卷
|
2卷引用:2024届安徽省芜湖市镜湖区安徽师范大学附属中学高三下学期5月最后一卷生物试题
解题方法
9 . 油菜素内脂(BR)是一种植物激素,它与IAA均能促进胚芽鞘的生长。为了研究BR及其与IAA之间的关系,科学家以大麦胚芽鞘为材料进行了相关实验,表2是在进行了预实验确定了IAA和BR最适浓度之后的正式实验,实验结果如表所示,下列叙述不正确的是( )
表1
表2
(注:IAA 1h→BR表示用IAA处理1h后用BR处理:BR lh→IAA表示用BR处理1h后用IAA处理。)
编号 | 1 | 2 | 3 | 4 | 5 | 6 |
BR的浓度/(mg·L-1) | 0 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 |
胚芽鞘的平均长度/(cm) | 16 | 20 | 38 | 51 | 42 | 20 |
组别 | 1 | 2 | 3 | 4 | 5 | 6 |
处理 | 蒸馏水 | IAA | BR | IAA 1h→BR | IAA+BR | BR 1h→IAA |
伸长量/cm | 0.71 | 1.04 | 1.12 | 1.15 | 1.60 | 1.53 |
(注:IAA 1h→BR表示用IAA处理1h后用BR处理:BR lh→IAA表示用BR处理1h后用IAA处理。)
| A.表中数据显示BR对株高的调节作用具有两重性 |
| B.促进幼苗生长的最适BR浓度在0.3-0.4mgL-1之间 |
| C.表中数据表明激素使用顺序不同对胚芽鞘生长的促进效果不同 |
| D.BR和IAA同时处理或先施用BR时,两者能表现出明显的协同作用 |
您最近一年使用:0次
2024-02-23更新
|
107次组卷
|
2卷引用:江西省上饶市2023-2024学年高三下学期第一次高考模拟考试生物试题
10 . 大豆是中国重要粮食作物之一,已有五千年栽培历史,古称“菽”。
(1)大豆种子中富含蛋白质,可用_______对其蛋白质含量进行定量测定。
(2)大豆种子萌发时,呼吸作用强度变化如图,下列分析正确的是________
在大豆种植中,种植行距与产量密切相关,种子粒重是决定大豆产量的关键性状之一。为探究种植行距对产量的影响,科学家进行了以下实验,实验处理和结果如表。
(3)去除顶芽可以提高主茎有效分枝数。施加下列______ 物质可以达到相同效果?(编号选填)
①生长素②2,4-D ③细胞分裂素④乙烯⑤脱落酸
(4)下列对上述实验结果的描述恰当的是________
(5)请结合表,说明如何能提高种子粒重?并阐述其能提高的原因。_____________ 。
大豆是一种光周期敏感的作物,其生育期和产量受光周期变化的影响非常大。近来,我国科学家研究发现了能对光周期进行响应从而控制种子粒重的关键基因Dt1。
(7)图中的Gm蛋白可将蔗糖从种皮运输到胚,促进胚的生长发育。据图简述短日照和长日照对大豆产量影响的机制。_____________________________ 。
(8)光周期除对种子粒重有影响外,光作为刺激信号,对植物造成的影响还可能有________
(9)结合上述信息和所学知识,下列提高大豆产量的建议合理的是________
(1)大豆种子中富含蛋白质,可用_______对其蛋白质含量进行定量测定。
| A.分光光度法 | B.同位素标记法 | C.引流法 | D.层析法 |
(2)大豆种子萌发时,呼吸作用强度变化如图,下列分析正确的是________
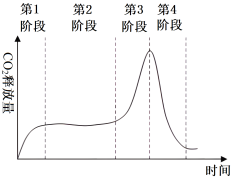
| A.第1阶段的种子中大量葡萄糖进入线粒体 |
| B.第2阶段的种子中产生大量乳酸 |
| C.第3阶段的种子温度高于第1和第2阶段 |
| D.第4阶段的种子呼吸速率下降可能是由于营养物质被消耗 |
在大豆种植中,种植行距与产量密切相关,种子粒重是决定大豆产量的关键性状之一。为探究种植行距对产量的影响,科学家进行了以下实验,实验处理和结果如表。
组别 | 种植行距(cm) | 株高(cm) | 主茎有效分枝数(个) | 百粒重(g) |
第1组 | 70 | 68.7 | 4.2 | 41.2 |
第2组 | 45 | 70.2 | 3.9 | 40.6 |
第3组 | 30 | 73.3 | 3.5 | 39.7 |
(3)去除顶芽可以提高主茎有效分枝数。施加下列
①生长素②2,4-D ③细胞分裂素④乙烯⑤脱落酸
(4)下列对上述实验结果的描述恰当的是________
| A.扩大种植行距,种子粒重增大 | B.株高越高,种子粒重越大 |
| C.种植行距与种子粒重无关 | D.主茎有效分枝数与种子粒重无关 |
(5)请结合表,说明如何能提高种子粒重?并阐述其能提高的原因。
大豆是一种光周期敏感的作物,其生育期和产量受光周期变化的影响非常大。近来,我国科学家研究发现了能对光周期进行响应从而控制种子粒重的关键基因Dt1。
| A.细胞质基质 | B.类囊体膜 | C.叶绿体基质 | D.细胞壁 |
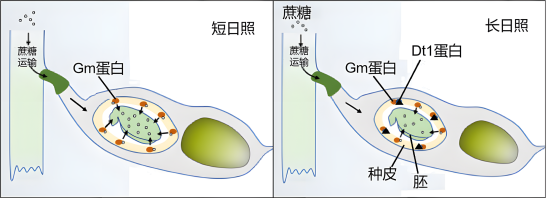
(7)图中的Gm蛋白可将蔗糖从种皮运输到胚,促进胚的生长发育。据图简述短日照和长日照对大豆产量影响的机制。
(8)光周期除对种子粒重有影响外,光作为刺激信号,对植物造成的影响还可能有________
| A.决定植物生长的方向 | B.影响种子的萌发 |
| C.诱导叶绿素形成 | D.影响植物的生长 |
(9)结合上述信息和所学知识,下列提高大豆产量的建议合理的是________
| A.种植行距尽可能大 | B.适当提高昼夜温差 |
| C.种子发育时,尽可能采用短日照 | D.适当提高二氧化碳浓度 |
您最近一年使用:0次
2024-04-12更新
|
171次组卷
|
3卷引用:2024届上海市徐汇区高三二模生物试卷


