(1)玉米茎的高度有高茎和矮茎两种类型,A控制高茎,将高茎植株与矮茎植株杂交,F1均为高茎,F1自交所得F2中高茎与矮茎的比例为3:1。在连续繁殖高茎玉米品系的过程中,偶然发现一株矮茎玉米突变体M,M自交,子代中高茎玉米植株占1/4。研究发现,M是由亲代高茎玉米植株一个A基因突变而成,基因型表示为AA+。让M自交,后代出现高茎玉米和矮茎玉米,利用遗传定律的实质解释产生此现象的原因:
(2)玉米的大穗杂种优势性状由两对等位基因(B1、B2和C1、C2)共同控制,两对等位基因都纯合时表现为衰退的小穗性状。若基因型为B1B2C1C2的大穗杂交种自交后代出现衰退小穗性状的概率为1/2,则说明
(3)玉米籽粒胚乳的颜色有黄色、紫色和杂色,科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制。已知胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。研究人员为研究胚乳颜色形成的机制,所做杂交实验及结果如下表所示,并作出如下推测:
表1
杂交组合 | F1胚乳颜色 | |
一 | 紫色RR(♀)×黄色rr(♂) | 紫色 |
二 | 紫色RR(♂)×黄色rr(♀) | 杂色 |
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体N所做杂交实验及结果如下表所示。
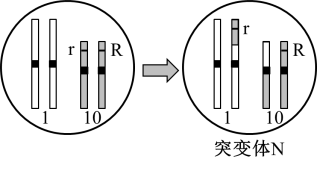
表2
杂交组合 | 部分F1胚乳 | ||
基因型 | 颜色 | ||
Rrr | 杂色 | ||
三 | 野生型rr(♀)x突变体N Rr(♂) | RRrr | 杂色 |
四 | 野生型rr(♂)x突变体N Rr(♀) | RRr | 紫色 |
(1)胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。从染色体组成分析,胚乳属于
杂交组合 | F1胚乳颜色 | |
一 | 紫色RR(♀)×黄色rr(♂) | 紫色 |
二 | 紫色RR(♂)×黄色rr(♀) | 杂色 |
推测一:可能与胚乳中R基因的数量有关;推测二:可能与胚乳中R基因的来源有关。
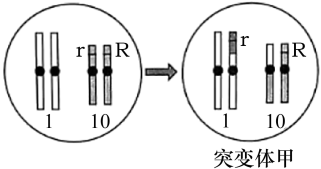
(2)为证实上述推测,研究人员利用突变体甲进行了相关实验。
杂交组合 | 部分F1胚乳 | ||
基因型 | 颜色 | ||
三 | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
四 | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
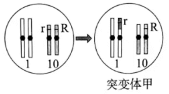
①突变体甲的形成过程如上图,形成甲的过程中发生的变异类型是
②研究发现,甲在产生配子时,10号染色体分离有时发生异常,但不影响配子的育性。表中F1出现少量基因型为RRrr的胚乳的原因是:突变体甲产生配子时,
③表中杂交结果仅支持推测
(3)研究人员推测在雌配子形成过程中,M基因表达产物可以降低R基因甲基化水平,使其表达不被抑制。现有M基因纯合突变体乙(mmRR)、野生型紫色玉米(MMRR)和黄色玉米(MMrr)。欲通过杂交实验验证上述推测,请写出实验方案并预期结果。
实验组:
对照组:
第一项实验:果蝇长翅(A)对残翅(a)为显性,红眼(B)对白眼(b)为显性。现有两组杂交实验,结果如下。

(1)若实验一为正交,则实验二为
(2)实验一的F1雌雄个体间相互交配,预期F2代基因型共有
第二项实验:果蝇中红眼(B)对白眼(b)为显性。有科学家进行下列实验,发现在F1中出现了1只例外的白眼雌蝇,请分析:
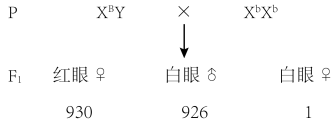
(3)若该白眼♀果蝇的产生来自性染色体的数目变异,下表是果蝇性染色体变异及对应性状。该白眼♀果蝇的基因型为
| XXY | 雌性可育 |
| XYY | 雄性可育 |
| XXX、YO、YY | 胚胎期死亡 |
| XO | 雄性不育 |
注:表中XO、YO分别表示只有一条X染色体、只有一条Y染色体
(4)检验该果蝇产生的原因,可用
(1)玉米的大穗杂种优势性状由两对等位基因(B1B2C1C2)共同控制,两对等位基因都纯合时表现为衰退的小穗性状。若大穗杂交种(B1B2C1C2)自交后代出现衰退的小穗性状的概率为1/2,则说明
白粉病是一种造成玉米严重减产的病害。科研人员利用基因工程技术将抗白粉病基因随机转入玉米中,获得抗病株。为进一步研究抗病株的遗传特性,让抗病株与原种进行杂交,结果如表所示:
组别 | 亲本 | F1 | F2表型及比例 |
1 | ♀抗病株×♂原种 | 自交 | 原种:抗病株≈4.93:1 |
2 | ♂抗病株×♀原种 | 自交 | 原种:抗病株≈4.85:1 |
(2)由此可知,相对于原种的感病性状,抗病性状为
(3)玉米籽粒胚乳的颜色有黄色、紫色和杂色,科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制。已知胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。胚乳是含有一整套精子染色体的三倍体。
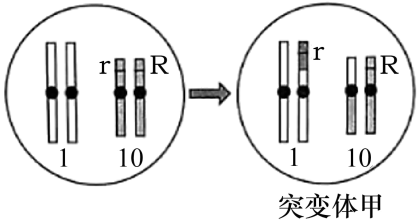
表1
杂交组合 | F1胚乳颜色 | |
一 | 紫色RR(♀)×黄色rr(♂) | 紫色 |
二 | 紫色RR(♂)×黄色rr(♀) | 杂色 |
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。
表2
杂交组合 | 部分F1胚乳 | ||
基因型 | 颜色 | ||
三 | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
四 | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
(1)玉米的大穗杂种优势性状由两对等位基因(B1B2C1C2)共同控制,两对等位基因都纯合时表现为衰退的小穗性状。若大穗杂交种(B1B2C1C2)自交后代出现衰退的小穗性状的概率为1/2,则说明
白粉病是一种造成玉米严重减产的病害。 科研人员利用基因工程技术将抗白粉病基因随机转入玉米中,获得抗病株。为进一步研究抗病株的遗传特性,让抗病株与原种进行杂交,结果如表所示:
组别 | 亲本 | F1 | F2表型及比例 |
1 | ♀抗病株x ♂原种 | 自交 | 原种:抗病株≈4.93:1 |
2 | ♂抗病株x♀原种 | 自交 | 原种:抗病株≈4.85:1 |
(3)玉米籽粒胚乳的颜色有黄色、紫色和杂色,科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制。已知胚乳是由精子与母本产生的两个极核融合后发育而成,每个极核的染色体组成均与卵细胞一致。胚乳是含有一整套精子染色体的三倍体。
杂交组合 | F1胚乳颜色 | |
一 | 紫色RR(♀)×黄色rr(♂) | 紫色 |
二 | 紫色RR(♂)×黄色rr(♀) | 杂色 |
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。
表2
杂交组合 | 部分F1胚乳 | ||
基因型 | 颜色 | ||
三 | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
四 | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
(1)果蝇的正常翅(A)对小翅(a)为显性。一只正常翅雌果蝇与另一只正常翅雄果蝇杂交,子代雄果蝇一半为正常翅、一半为小翅,雌果蝇全部为正常翅。则控制果蝇翅形的基因A、a位于
(2)果蝇的正常翅与残翅由2号染色体上的B、b基因控制。纯合的正常翅果蝇与残翅果蝇杂交子代中除一只果蝇为残翅外其余均为正常翅,这对相对性状中,隐性性状是
实验步骤:
①用该残翅果蝇与纯合正常翅果蝇杂交,获得F1;
②F1雌雄果蝇自由交配,观察并统计F2表型及比例。
预期实验结果及实验结论:
Ⅰ.若F2表型及比例为
Ⅱ.若F2表型及比例为
(3)果蝇的灰体与黑体是一对相对性状,由D、d基因控制。现将一只正常翅灰体果蝇与一只残翅黑体果蝇杂交,F1全部为正常翅灰体,F1与残翅黑体果蝇杂交实验结果如下表所示。
杂交组合 | 父本 | 母本 | F2表型及比例 |
一 | F1 | 残翅黑体 | 1正常翅灰体∶1残翅黑体 |
二 | 残翅黑体 | F1 | 5正常翅灰体∶1正常翅黑体∶1残翅灰体∶5残翅黑体 |
由表中数据可知,杂交组合一与组合二的F2表型及比例不同的原因是
长翅红眼 | 长翅白眼 | 残翅红眼 | 残翅白眼 | |
雌蝇 | 3/8 | 0 | 1/8 | 0 |
雄蝇 | 3/16 | 3/16 | 1/16 | 1/16 |
(2)该果蝇与“另一亲本”杂交,子代的长翅红眼雌果蝇中纯合体占的比例为
(3)若只根据子代果蝇的眼色就能判断其性别,则选作亲本果蝇的眼色基因型为
(4)若同时考虑上述两对性状,子代雌雄果蝇都有四种表现型,且比例为3:3:1:1,则选作亲本果蝇的基因型雌雄分别为
(5)用残翅白眼雌果蝇(aaXbXb)与长翅红眼雄果蝇(AAXBY)杂交,F1雌果蝇表现为长翅红眼,雄果蝇表现为长翅白眼。从F2长翅红眼雌果蝇和长翅白眼雄果蝇中各随机选取一只杂交,子代中出现残翅白眼果蝇的概率为
(1)根据染色体的组成判断,胚乳属于 。
| A.单倍体 | B.二倍体 | C.三倍体 | D.四倍体 |
(2)科研人员进行了系列实验来研究胚乳颜色形成的遗传学机制:
| 杂交组合 | F1胚乳颜色 | |
| I | 紫色RR(♀)×黄色rr(♂) | 紫色 |
| Ⅱ | 紫色RR(♂)×黄色rr(♀) | 杂色 |
①RRR ②RRr ③Rrr ④mr ⑤RRrr ⑥Rrrr
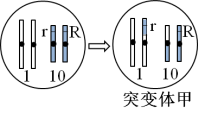
研究人员对胚乳颜色形成的机制作出如下推测。推测一:可能与胚乳中R基因的数量有关;推测二:可能与胚乳中R基因的来源有关。为证实上述推测,研究人员利用突变体甲进行了相关实验(如表)。
杂交组合 | 部分F1胚乳 | ||
| 基因型 | 颜色 | ||
| Ⅲ | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
| RRrr | 杂色 | ||
| IV | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |
(3)如图所示,突变体甲的获得所依据的原理是
| A.基因重组 | B.基因突变 | C.染色体数目变异 | D.染色体结构变异 |
(4)研究发现,突变体甲在产生配子时,10号染色体分离有时发生异常,但不影响配子的育性。表中基因型为RRrr的胚乳产生的原因是:突变体甲产生配子时,___________。
| A.减I后期10号染色体的姐妹染色单体未分开 |
| B.减II后期10号染色体的姐妹染色单体未分开 |
| C.减I后期10号染色体的同源染色体未分离 |
| D.减II后期10号染色体的同源染色体未分离 |
(5)上表杂交结果仅支持推测
(6)在第(5)题结论的基础上,研究人员推测在雌配子形成过程中,M基因表达产物可以降低R基因甲基化水平,使其表达不被抑制。现有M基因纯合突变体乙(mmRR)、野生型紫色玉米(MMRR)和黄色玉米(MMrr)。欲通过杂交实验验证该推测,(1)一组杂交方案为
①突变体乙(mmRR)做母本与黄色玉米(MMrr)做父本杂交
②黄色玉米(MMrr)做母本与突变体乙(mmRR)做父本杂交
③野生型紫色玉米(MMRR)做母本与黄色玉米(MMrr)做父本杂交
④黄色玉米(MMrr)做母本与野生型紫色玉米(MMRR)做父本杂交
⑤子代胚乳表现为紫色
⑥子代胚乳表现为杂色
子一代褐色(只) | 子一代灰色(只) | 子一代黄色(只) | 子一代白色(只) | |
单对杂交一 | 10 | 15 | 15 | 5 |
单对杂交二 | 19 | 31 | 30 | 9 |
单对杂交三 | 11 | 14 | 15 | 6 |
总计 | 40 | 60 | 60 | 20 |
| A.杂交组合一的亲本基因型为AaBb×AaBb |
| B.该动物毛色遗传规律的本质为非同源染色体上的非等位基因自由组合 |
| C.子一代出现上表4种表现型比例的原因可能是基因型为AB的配子致死 |
| D.若杂交组合二中亲本减数分裂过程中发生互换,则子一代毛色的性状分离比会发生变化 |

(1)正常基因型为Rr的玉米进行自交,获得的玉米籽粒胚乳的基因组成有
(2)紫色RR(♂)×黄色rr(♀),F1胚乳为紫色,反交时F1胚乳为杂色。对胚乳颜色形成的机制有人作出了以下两种推测。
推测一:可能与胚乳中R基因的数量有关;
推测二:可能与胚乳中R基因的来源有关。
为证实上述推测,研究人员利用突变体甲进行了相关实验。结果如下表。
杂交组合 | 部分F1胚乳 | ||
基因组成 | 颜色 | ||
I | 野生型rr(♀)×甲Rr(♂) | Rrr | 杂色 |
RRrr | 杂色 | ||
II | 野生型rr(♂)×甲Rr(♀) | RRr | 紫色 |


