| 处理时间 | GA3浓度(μg·mL-1) | ||||
| (h) | 0 | 25 | 50 | 75 | 100 |
| 2 | 11 | 11.2 | 11.3 | 30 | 39 |
| 4 | 11 | 11.7 | 16.7 | 34 | 56.3 |
| 6 | 11 | 13.3 | 32.3 | 40.3 | 77.3 |
| 8 | 11 | 19.3 | 36.7 | 63.7 | 87 |
| A.尽管薰衣草体内的赤霉素只有一种,含量微少,但起重要的调节作用 |
| B.相同处理时间下,随着赤霉素浓度的增加,种子的发芽率有所增加 |
| C.相同赤霉素浓度下,随着处理时间的增加,发芽率呈逐渐升高的趋势 |
| D.为有效促进薰衣草种子的萌发,以100μgmL-1的赤霉素处理8小时左右为佳 |
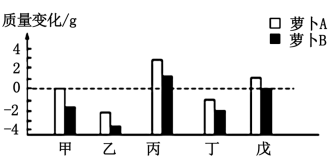
| A.实验开始时,萝卜A比B的细胞液浓度高 |
| B.甲~戊组中,蔗糖溶液浓度最大的是乙组 |
| C.萝卜A和B均未发生质壁分离的是乙组和丁组 |
| D.戊组中,萝卜B与蔗糖溶液的水分交换处于动态平衡 |
试管1 | 试管2 | 试管3 | 试管4 | |
底物 | 5cm3的蛋白块 | 5cm3的蛋白块 | 5cm3的蛋白块 | 5cm3的蛋白块 |
试剂 | 4mL蒸馏水 | ① | 4mL胶原蛋 白酶溶液 | 4mL胶原蛋 白酶溶液 |
温度 | 37℃水浴 | 37℃水浴 | 37℃水浴 | 0℃水浴 |
pH | pH=7 | pH=7 | pH=1 | ② |
实验现象 | 蛋白块无 明显变化 | 蛋白块 明显减小 | 蛋白块 无明显变化 | 蛋白块无 明显变化 |
| A.选择蛋白块为底物的原因是蛋白块较容易观察 |
| B.表中①②分别为4mL胶原蛋白酶溶液、pH=7 |
| C.单位时间蛋白块体积的变化可反映胶原蛋白酶活性大小 |
| D.试管3实验一段时间后,将其pH上调至7,一段时间后发现蛋白块大小会明显减小 |
| A.本实验的目的是探究盆栽黄瓜幼苗叶片发黄是由缺 N还是缺 Mg引起叶绿素合成不足导致的 |
| B.若A、C两组叶片变绿,B组叶片发黄,则说明该批盆栽黄瓜幼苗叶片发黄是由于缺N造成的 |
| C.若A 组叶片发黄,B、C 两组叶片变绿,则说明该批盆栽黄瓜幼苗叶片发黄是由于缺 Mg造成的 |
| D.若A、B两组叶片发黄,C组叶片变绿,则说明该批盆栽黄瓜幼苗叶片发黄是由于缺少N或 Mg造成的 |
不同离子浓度对解脂酶活性的影响
表1相对低浓度
离子类别 | 离子浓度/(mol/L) | |||||
0 | 0.002 | 0.004 | 0.006 | 0.008 | 0.01 | |
Ca2+ | 0.518 | 0.520 | 0.513 | 0.476 | 0.458 | 0.430 |
Ba2+ | 0.518 | 0.509 | 0.479 | 0.452 | 0.417 | 0.361 |
Cu2+ | 0.518 | 0.492 | 0.467 | 0.416 | 0.362 | 0.319 |
Zn2+ | 0.518 | 0.532 | 0.573 | 0.615 | 0.671 | 0.735 |
Fe2+ | 0.518 | 0.535 | 0.562 | 0.594 | 0.618 | 0.632 |
Mg2+ | 0.518 | 0.542 | 0.580 | 0.646 | 0.719 | 0.797 |
离子类别 | 离子浓度/(mol/L) | ||||
0.02 | 0.04 | 0.06 | 0.08 | 0.1 | |
Ca2+ | 0.406 | 0.356 | 0.316 | 0.249 | 0.167 |
Ba2+ | 0.318 | 0.236 | 0.187 | 0.125 | 0.051 |
Cu2+ | 0.224 | 0.106 | 0.056 | 0.038 | 0.03 |
Zn2+ | 0.719 | 0.635 | 0.503 | 0.329 | 0.123 |
Fe2+ | 0.650 | 0.589 | 0.498 | 0.371 | 0.213 |
Mg2+ | 0.781 | 0.743 | 0.684 | 0.619 | 0.516 |
| A.实验中可以利用氯化钙、硫酸铜溶液分别提供钙离子和铜离子 |
| B.实验自变量为离子种类和离子浓度,该解脂酶的活性为因变量 |
| C.在相对低浓度下随离子浓度增加,Ca2+、Ba2+、Cu2+会使该酶的活性降低 |
| D.Zn2+、Fe2+、Mg2+三种金属离子在相对高浓度下会使酶空间结构改变 |

| A.JA和乙烯作为信号分子通过调控基因表达影响植物对盐胁迫的抵抗能力 |
| B.与NaCl组相比,加入乙烯合成抑制剂、JA合成抑制剂组的胚根长度均明显增加 |
| C.由实验结果可知,在盐胁迫条件下JA和乙烯对水稻胚根生长均有促进作用 |
| D.欲探究JA与乙烯之间的作用关系,还需在此实验基础上再设置一组实验 |
| 实验 | 实验组的处理 | 实验结果 |
| 第一组 | 在外界葡萄糖浓度为5mmol/L时,用蛋白质合成抑制剂处理小肠微绒毛,一段时间后测量其葡萄糖的转运速率 | 对照组葡萄糖的转运速率>实验组葡萄糖的转运速率,且实验组葡萄糖的转运速率>0 |
| 第二组 | 在外界葡萄糖浓度为5mmol/L时,用呼吸抑制剂处理小肠微绒毛,一段时间后测量其葡萄糖的转运速率 | 对照组葡萄糖的转运速率>实验组葡萄糖的转运速率,且实验组葡萄糖的转运速率=0 |
| A.对照组需在外界葡萄糖浓度为5mmol/L的条件下进行 |
| B.蛋白质合成抑制剂会对膜上载体蛋白的功能产生抑制 |
| C.呼吸抑制剂会对细胞的无氧呼吸和有氧呼吸产生抑制 |
| D.该实验证明小肠黏膜上皮细胞吸收葡萄糖的过程为主动运输 |

| A.对照组为空白对照,主茎顶端不放置琼脂块 |
| B.GR24与NAA对侧枝的生长均有抑制作用 |
| C.拟南芥植株上正常生长的侧枝长度小于对照组 |
| D.GR24组侧枝长度小于对照组可能是由于GR24抑制了侧芽处生长素向外运输 |
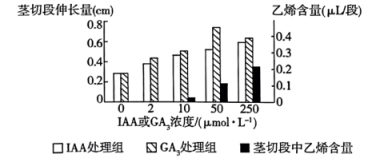
| A.本实验的自变量为植物激素种类和浓度,培养时间是无关变量 |
| B.GA3促进菟丝子茎切段伸长的最适浓度在10~250μmol·L-1范围内 |
| C.空白对照组中茎切段缓慢生长是因为存在少量内源激素 |
| D.据图可知,生长素通过促进乙烯的合成来促进植物的生长 |
不同浓度IAA、GA3、CTK处理番茄种子的发芽率和发芽势
生长调节剂种类 | IAA | GA3 | CTK | ||||||
浓度(mg/L) | 发芽势(%) | 发芽率(%) | 浓度(mg/L) | 发芽势(%) | 发芽率(%) | 浓度(mg/L) | 发芽势(%) | 发芽率(%) | |
检测指标 | 2 | 91.00 | 96.13 | 100 | 94.00 | 97.93 | 10 | 94.00 | 97.93 |
4 | 89.00 | 93.13 | 200 | 89.00 | 94.20 | 25 | 90.00 | 95.20 | |
6 | 85.00 | 89.46 | 300 | 84.00 | 91.60 | 50 | 87.00 | 91.86 | |
8 | 80.00 | 84.06 | 400 | 81.00 | 88.40 | 100 | 84.00 | 86.60 | |
10 | 76.00 | 81.53 | 500 | 68.00 | 74.00 | 200 | 80.00 | 82.60 | |
| A.实验所用番茄种子形态、大小等一致,对照组番茄种子用蒸馏水处理 |
| B.由表可知,GA3和IAA对番茄种子发芽势均表现为低浓度促进,高浓度抑制 |
| C.由表可知,当GA3浓度为500mg/L时,番茄种子发芽率低于CK,较CK组降低6% |
| D.由表可知,一定范围内随着CTK浓度的升高,番茄种子发芽势和发芽率都逐渐降低 |


