名校
1 . 某雌雄异株植物(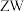 )的红果和黄果由基因
)的红果和黄果由基因 控制,叶片菱形和卵形由基因
控制,叶片菱形和卵形由基因 控制,两对基因独立遗传,实验人员选择红果菱形雄株(甲)、红果卵形雌株(乙)、黄果菱形雌株(丙)进行了下表所示实验。不考虑
控制,两对基因独立遗传,实验人员选择红果菱形雄株(甲)、红果卵形雌株(乙)、黄果菱形雌株(丙)进行了下表所示实验。不考虑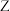 、
、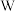 染色体的同源区段,回答下列问题:
染色体的同源区段,回答下列问题:
(1)根据实验二结果分析,两对等位基因中位于染色体上的是基因________ ,判断的依据是________________ 。
(2)实验一中, 全为雌株的原因可能是
全为雌株的原因可能是________________ ,植株甲的基因型是________ 。
(3)取实验一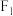 中卵形叶雌株并对其进行射线处理,进行培育后与杂合的菱形叶雄株杂交,分别统计
中卵形叶雌株并对其进行射线处理,进行培育后与杂合的菱形叶雄株杂交,分别统计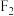 单株的表型及比例,发现其中一个株系的
单株的表型及比例,发现其中一个株系的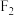 中雌、雄株都表现为
中雌、雄株都表现为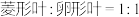 。已确定该结果的出现与同源染色体片段转移有关。
。已确定该结果的出现与同源染色体片段转移有关。
①发生的具体同源染色体片段转移是________ ,且需要满足的条件是________________ (从配子的可育性方面作答)。
②若让该株系的菱形叶雌、雄株杂交,则其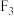 的表型及比例是
的表型及比例是________ (写出性别)。
③为验证该染色体片段转移,实验人员选择该株系中________ (填“雄株”、“雌株”或“雄株和雌株”)与正常的雌、雄植株细胞制成临时装片,在显微镜下观察并比较它们的性染色体结构。与正常植株相比,若________________ ,则可初步证明该株系确实发生了染色体片段转移。
)的红果和黄果由基因控制,叶片菱形和卵形由基因控制,两对基因独立遗传,实验人员选择红果菱形雄株(甲)、红果卵形雌株(乙)、黄果菱形雌株(丙)进行了下表所示实验。不考虑、染色体的同源区段,回答下列问题:实验 | 亲本 |
|
一 | 甲 | 红果菱形雌株:红果卵形雌株:黄果菱形雌株:黄果卵形雌株=3:3:1:1 |
二 | 甲 | 红果菱形雄株:黄果菱形雄株:红果菱形雌株:红果卵形雌株:黄果菱形雌株:黄果卵形雌株=2:2:1:1:1:1 |
 乙染色体上的是基因
乙染色体上的是基因(2)实验一中,
全为雌株的原因可能是(3)取实验一
中卵形叶雌株并对其进行射线处理,进行培育后与杂合的菱形叶雄株杂交,分别统计单株的表型及比例,发现其中一个株系的中雌、雄株都表现为。已确定该结果的出现与同源染色体片段转移有关。①发生的具体同源染色体片段转移是
②若让该株系
的菱形叶雌、雄株杂交,则其的表型及比例是③为验证该染色体片段转移,实验人员选择该株系
中
您最近一年使用:0次
2023-04-24更新
|
1291次组卷
|
5卷引用:2023届广东省湛江市高三二模生物试题
2023届广东省湛江市高三二模生物试题(已下线)生物(广东卷)-学易金卷:2023年高考考前押题密卷(含考试版、全解全析、参考答案、答题卡)2023届广东省广州大学附中高三三模生物试题湖南省部分学校2022-2023学年高三4月月考生物试题2023届辽宁省辽阳市高三二模生物试题
名校
解题方法
2 . 雌性哺乳动物在早期胚胎发育过程中 1 条 X 染色体会发生整体范围内的沉默(“非逃逸”基因转录活性被抑制),X 染色体失活(XCI),仅有少数“逃逸”基因仍能正常表达。XCI能保证雌、雄间 X 染色体上基因表达水平的一致,也会造成体外受精胎儿性别比例的失衡。
(1)为制备体外受精(IVF)的胚胎,研究人员采用超数排卵方法采集小鼠______ 期卵母细胞,另一方面,将成年健康公鼠的精子在体外条件下进行______ 处理,将二者置于适当的培养液中完成体外受精。收集早期胚胎,移植到经过______ 处理的代孕母鼠中,获得的胚胎含有______ (填“单”“双”或“三”)亲的遗传物质。
(2)研究人员对体外受精胎儿性别进行鉴定,表中数据显示正常胎儿性别比例(雄/雌)______ 自然生产(IVO)胎儿的性别比例。
(3)为进一步揭示体外受精的胎儿性别比例失衡的原因,研究人员剖取第 7.5 天的小鼠胚胎进行胎儿发育情况统计,结果如图 1:__________________ 。
②研究人员收集不同组别的胚胎细胞,定量检测具代表性的“非逃逸”基因,结果显示这些基因的表达在 IVF 雌性胚胎中显著______ (填“低于”“等于”或“高于”)IVO 雌性胚胎,说明 IVF雌性胚胎 XCI 不足。
③已有研究表明,XCI 高度依赖激活因子 RNF12,研究人员利用免疫荧光技术检测 RNF12 蛋白的表达量如图 2,结果为________________________ 。________________________ 。
(1)为制备体外受精(IVF)的胚胎,研究人员采用超数排卵方法采集小鼠
(2)研究人员对体外受精胎儿性别进行鉴定,表中数据显示正常胎儿性别比例(雄/雌)
组别 | 胚胎数 | 活仔率(%) | 雄/雌 |
自然生产(IVO) | 81 | 56.25 | 1.08 |
体外受精(IVF) | 82 | 45.58 | 1.48 |
(3)为进一步揭示体外受精的胎儿性别比例失衡的原因,研究人员剖取第 7.5 天的小鼠胚胎进行胎儿发育情况统计,结果如图 1:

②研究人员收集不同组别的胚胎细胞,定量检测具代表性的“非逃逸”基因,结果显示这些基因的表达在 IVF 雌性胚胎中显著
③已有研究表明,XCI 高度依赖激活因子 RNF12,研究人员利用免疫荧光技术检测 RNF12 蛋白的表达量如图 2,结果为

您最近一年使用:0次
2024-05-14更新
|
80次组卷
|
5卷引用:广东实验中学2023-2024学年高二下学期期中考试生物试题
3 . 现有一未受人类干扰的自然湖泊,研究小组调查了该湖泊中某食物链最高营养级的某种鱼种群的年龄组成,结果如下表。
注:表中“1+”表示鱼的年龄≥1且<2,其他以此类推;该鱼在3十时达到性成熟(进入成年),9十时丧失繁殖能力(进入老年)请回答下列相关问题:
(1)根据表中数据可知,该鱼群的年龄结构大体为__________ ,理由是__________ ,一段时间内,该鱼种群数量的变化趋势是__________ 。
(2)如果要调查该鱼的种群密度,常用的调查方法是标记重捕法。该方法常用于调查__________ (填特点)动物的种群密度。
(3)在自然湖泊中,该鱼种群的__________ 特征直接决定其种群密度,影响该鱼种群数量变化的非密度制约因素有__________ (答出2项即可)。
年龄 | 0+ | 1+ | 2+ | 3+ | 4+ | 5+ | 6+ | 7+ | 8+ | 9+ | 10+ | 11+ | ≥12 |
个体数 | 92 | 187 | 121 | 70 | 69 | 62 | 63 | 72 | 64 | 55 | 42 | 39 | 264 |
(1)根据表中数据可知,该鱼群的年龄结构大体为
(2)如果要调查该鱼的种群密度,常用的调查方法是标记重捕法。该方法常用于调查
(3)在自然湖泊中,该鱼种群的
您最近一年使用:0次
2023-07-20更新
|
192次组卷
|
4卷引用:广东省深圳市光明区2022-2023学年高二2月期末生物试题
真题
名校
4 . 回答下列果蝇眼色的遗传问题。
(1)有人从野生型红眼果蝇中偶然发现一只朱砂眼雄蝇,用该果蝇与一只红眼雌蝇杂交得F1,F1随机交配得F2,子代表现型及比例如下(基因用B、b 表示):
①B、b 基因位于_____ 染色体上,朱砂眼对红眼为_____ 性。
②让F2 代红眼雌蝇与朱砂眼雄蝇随机交配,所得F3代中,雌蝇有_____ 种基因型,雄蝇中朱砂眼果蝇所占比例为_____ 。
(2)在实验一F3的后代中,偶然发现一只白眼雌蝇。研究发现,白眼的出现与常染色体上的基用E 、e 有关。将该白眼雌蝇与一只野生型红眼雄蝇杂交得F’1,F’1随机交配得F’2, 子代表现型及比例如下:
实验二中亲本白眼雌蝇的基因型为_____ ;F’ 2 代杂合雌蝇共有_____ 种基因型,这些杂合雌蝇中红眼果蝇所占的比例为_____ 。
(3)果蝇出现白眼是基因突变导致的,该基因突变前的部分序列(含起始密码信息) 如下图所示。
(注:起始密码子为AUG,终止密码子为UAA,UAG 或UGA )

上图所示的基因片段在转录时,以_____ 链为模板合成mRNA ;若“↑”所指碱基对缺失,该基因控制合成的肽链含_____ 个氨基酸。
(1)有人从野生型红眼果蝇中偶然发现一只朱砂眼雄蝇,用该果蝇与一只红眼雌蝇杂交得F1,F1随机交配得F2,子代表现型及比例如下(基因用B、b 表示):
| 实验一 | 亲本 | F1 | F2 | ||
| 雌 | 雄 | 雌 | 雄 | ||
| 红眼(♀)×朱砂眼(♂ ) | 全红眼 | 全红眼 | 红眼:朱砂眼=1:1 | ||
②让F2 代红眼雌蝇与朱砂眼雄蝇随机交配,所得F3代中,雌蝇有
(2)在实验一F3的后代中,偶然发现一只白眼雌蝇。研究发现,白眼的出现与常染色体上的基用E 、e 有关。将该白眼雌蝇与一只野生型红眼雄蝇杂交得F’1,F’1随机交配得F’2, 子代表现型及比例如下:
| 实验二 | 亲本 | F’1 | F’2 | |
| 雌 | 雄 | 雌、雄均表现为 | ||
| 白眼(♀)×红眼(♂ ) | 全红眼 | 全朱砂眼 | 红眼 ︰朱砂眼 ︰白眼=4 ︰3 ︰1 | |
(3)果蝇出现白眼是基因突变导致的,该基因突变前的部分序列(含起始密码信息) 如下图所示。
(注:起始密码子为AUG,终止密码子为UAA,UAG 或UGA )
上图所示的基因片段在转录时,以
您最近一年使用:0次
2019-01-30更新
|
1756次组卷
|
12卷引用:2013-2014学年广东省广州市越秀区高一下学期期末水平调研测生物卷
(已下线)2013-2014学年广东省广州市越秀区高一下学期期末水平调研测生物卷2013年全国普通高等学校招生统一考试理科综合能力测试生物(四川卷)(已下线)2014届浙江诸暨牌头中学高三12月月考理综生物卷(已下线)2014年人教版高考生物二轮专题4.2遗传的基本规律和伴性遗传B(已下线)2013-2014学年黑龙江牡丹江一中高二上期期末考试生物卷(已下线)2015届高三生物一轮总复习备课资源包:专题10遗传的基本规律(已下线)2013-2014学年黑龙江牡丹江一中高二上期期中生物卷2015届浙江省宁波市鄞州高级中学高三第一次考试理科综合生物试卷2015-2016学年河南省洛阳一中高二上10月月考生物试卷2015-2016四川双流中学高二上学期入学考试生物试卷2015-2016学年吉林市一中高一下期末考试生物试卷2015-2016学年四川成都树德中学高一下期末考试生物试卷
名校
解题方法
5 . 不对称体细胞杂交是指利用射线破坏染色质的供体细胞,与未经射线照射的受体细胞融合,所得融合细胞含受体全部遗传物质及供体部分遗传物质。科研人员用大剂量的X射线处理拟南芥(2n=10)的原生质体,然后诱导其与柴胡(2n=12)的原生质体融合,最终获得不对称杂种植株。回答下列问题:
(1)为获得拟南芥和柴胡的原生质体,可将消毒后的外植体用小刀切碎,放入含有_____ 酶的缓冲液中,以获得原生质体。
(2)可采用二乙酸荧光素(FAD)法测定原生质体活力,已知FAD本身无荧光,当其进入细胞后可被酯酶分解为无毒、具有荧光的物质,该荧光物质不能透过细胞膜,会留在细胞内发出荧光。据此应选择_____ 的原生质体用于融合。
(3)获得的原生质体可使用_____ 等化学方法(写出一种)诱导融合,在杂种细胞形成杂种植株的过程中,需要避光培养的是_____ 阶段,诱导生根的过程中,细胞分裂素和生长素的比值_____ (“=1”、“>1”、“<1”)
(4)科研人员对获得的部分植株细胞进行染色体观察、计数和DNA分子标记鉴定,统计结果如下表所示:
由表可知,后代植株中_____ 类型一定不是杂种植株。科研人员推测杂种植株的染色体主要由柴胡亲本来源的染色体组成,拟南芥亲本的遗传物质可能不是以染色体的形式存在于杂种植株细胞中,而是以DNA片段的方式整合进柴胡的基因组,作出以上推测的依据是_____ 。
(1)为获得拟南芥和柴胡的原生质体,可将消毒后的外植体用小刀切碎,放入含有
(2)可采用二乙酸荧光素(FAD)法测定原生质体活力,已知FAD本身无荧光,当其进入细胞后可被酯酶分解为无毒、具有荧光的物质,该荧光物质不能透过细胞膜,会留在细胞内发出荧光。据此应选择
(3)获得的原生质体可使用
(4)科研人员对获得的部分植株细胞进行染色体观察、计数和DNA分子标记鉴定,统计结果如下表所示:
| 后代植株类型 | 染色体数目形态 | DNA分子标记鉴定 |
| 甲 | 12,与柴胡染色体相似 | 含双亲DNA片段 |
| 乙 | 12,与柴胡染色体相似 | 无拟南芥DNA片段 |
| 丙 | 12,与柴胡染色体相似 | 含双亲DNA片段和新的DNA片段 |
您最近一年使用:0次
7日内更新
|
27次组卷
|
2卷引用:广东省梅州市梅县东山中学2023-2024学年高二下学期期中考试生物试题
6 . 基因定位是遗传学研究的一项重要工作,基因定位的方法很多,请分析回答以下问题。
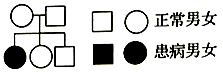
(1)系谱分析法能够判断基因位于性染色体或常染色体。如图为某单基因遗传病的系谱图,则该致病基因位于______________ 染色体上,判断的依据是______________ 。
(2)非整倍体测交法可以用来测定基因是否位于某条常染色体。已知植物的单体和三体产生的配子均为可育配子且后代均存活。如测定一对基因是否位于4号染色体上:
方法一: 用隐性突变型植株和野生型单体(4 号染色体少一条) 杂交,若子代中野生型和突变型植株之比是___________ 而不是1:0,则说明该突变基因及其等位基因位于4 号染色体上。
方法二: 用隐性突变型植株和野生型三体(4 号染色体多一条) 杂交,让子一代中的三体个体再和隐性亲本测交。请预测测交结果并得出相应结论:_______________________ 。
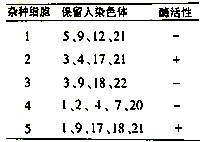
(3) 体细胞杂交法可将基因准确定位在某条染色体上。已知人的体细胞和小鼠的体细胞(2N=40) 杂交(融合)后,杂种细胞在有丝分裂过程中,人的染色体逐渐地丢失,最后仅保留少数几条。由于人染色体的丢失是随机的,因此不同的杂种细胞会保留不同的人染色体。如测定不同的杂种细胞中人胸苷激酶的活性,结果见下表。由表分析: 人胸苷激酶基因位于______________ 号染色体上。
(1)系谱分析法能够判断基因位于性染色体或常染色体。如图为某单基因遗传病的系谱图,则该致病基因位于
(2)非整倍体测交法可以用来测定基因是否位于某条常染色体。已知植物的单体和三体产生的配子均为可育配子且后代均存活。如测定一对基因是否位于4号染色体上:
方法一: 用隐性突变型植株和野生型单体(4 号染色体少一条) 杂交,若子代中野生型和突变型植株之比是
方法二: 用隐性突变型植株和野生型三体(4 号染色体多一条) 杂交,让子一代中的三体个体再和隐性亲本测交。请预测测交结果并得出相应结论:
(3) 体细胞杂交法可将基因准确定位在某条染色体上。已知人的体细胞和小鼠的体细胞(2N=40) 杂交(融合)后,杂种细胞在有丝分裂过程中,人的染色体逐渐地丢失,最后仅保留少数几条。由于人染色体的丢失是随机的,因此不同的杂种细胞会保留不同的人染色体。如测定不同的杂种细胞中人胸苷激酶的活性,结果见下表。由表分析: 人胸苷激酶基因位于
您最近一年使用:0次
解题方法
7 . 经济植物矮化突变体具有抗倒伏、适合密植等特点。下表是陆地棉矮化突变体与正常植株光合特性差异的比较。回答下列问题:
(1)叶绿素主要吸收_____ 光。若欲比较突变体和正常植株叶片中叶绿素含量的多少,可将提取的色素用层析液分离,分离的原理是_____ ,分离后观察滤纸条上第_____ 条色素带(从下往上计数)的宽窄并进行分析。
(2)根据光合作用的过程和表中数据分析可知,影响胞间CO2浓度的因素有_____ (答出2点即可)等方面。据表分析,与正常植株相比,现蕾后,矮化突变体净光合速率下降,原因可能是_____ 。
(3)据表推测,突变体矮化现象的出现,主要是在_____ (填“现蕾前”或“现蕾后”),理由是_____ 。
(4)矮化棉花由于适宜机械化生产,具有重要的应用价值,但其光合能力较正常植株弱,不利于光合产物积累,所以在生产中可以采取_____ (答出1点即可)措施增加其单位面积的光合产物积累量。
时期 | 材料 | 净光合速率/(μmol·m-2·s-1) | 气孔导度/(mmol·m-2·s-1) | 胞间CO2浓度/(μmol·mol-1) |
现蕾前 | 突变体 | 26.36 | 0.77 | 306.76 |
正常植株 | 26.74 | 0.85 | 314.58 | |
现蕾后 | 突变体 | 22.61 | 0.65 | 249.30 |
正常植株 | 25.75 | 1.01 | 261.89 |
(2)根据光合作用的过程和表中数据分析可知,影响胞间CO2浓度的因素有
(3)据表推测,突变体矮化现象的出现,主要是在
(4)矮化棉花由于适宜机械化生产,具有重要的应用价值,但其光合能力较正常植株弱,不利于光合产物积累,所以在生产中可以采取
您最近一年使用:0次
2024-06-13更新
|
50次组卷
|
3卷引用:广东省名校联盟2023-2024学年高二下学期期中质量检测生物试题
解题方法
8 . 科研人员为分析发生重度林火后的森林在不同演替阶段植物群落物种组成和多样性变化规律,选取4个不同演替阶段的植物群落,即火后演替5年、火后演替15年、火后演替23年和森林顶级群落作为一个演替序列,采用样方法调查植物群落的物种组成及个体数量,结果见下表。请回答下列问题:
表 不同演替阶段植物群落的物种组成及个体数量
(1)发生重度林火后的森林进行_________ 演替,做出该判断的理由是__________ 。
(2)随着演替的进行,灌木层植物个体的数量表现出________ 的变化趋势。乔木层形成后,草本植物的个体数出现急剧下降的原因是__________ 。
(3)发生重度林火后,不同生态位的植物能在短时期侵入并定居,植物物种数达到最高。但随着群落演替的进行,植物个体数量呈现_________ 趋势。从种间关系的角度分析其原因是__________ 。
(4)当群落演替到森林顶级群落后,在中等或高等强度的林火干扰后,群落的物种多样性会上升,原因是_____________ (答出一点即可)。
表 不同演替阶段植物群落的物种组成及个体数量
| 演替阶段 | 物种组成(种) | 个体数量/[株·(400m2)-1] | ||||||
| 森林顶级群落 | 乔木 | 灌木 | 草本 | 总计 | 乔木 | 灌木 | 草本 | 总计 |
| 火后演替5年 | — | 24 | 76 | 100 | — | 131 | 3579 | 3710 |
| 火后演替15年 | — | 17 | 46 | 63 | — | 196 | 2773 | 2969 |
| 火后演替23年 | 7 | 18 | 39 | 64 | 151 | 204 | 2077 | 2432 |
| 森林顶级群落 | 12 | 35 | 30 | 77 | 112 | 171 | 1476 | 1759 |
(2)随着演替的进行,灌木层植物个体的数量表现出
(3)发生重度林火后,不同生态位的植物能在短时期侵入并定居,植物物种数达到最高。但随着群落演替的进行,植物个体数量呈现
(4)当群落演替到森林顶级群落后,在中等或高等强度的林火干扰后,群落的物种多样性会上升,原因是
您最近一年使用:0次
21-22高二·全国·单元测试
名校
9 . 下图是缩手反射的反射弧结构模式图,请据图答题。________________ 。
(2)图甲中反射弧通常由①、②、③、④____________ 和⑤__________ 组成。当针刺手指时,兴奋在反射弧中的传递只能是____________ (填“单向”或“双向”)的。
(3)在未受刺激时,②细胞膜两侧的电位处于_________ 状态,表现为_______________ 。
(4)图中的③、④属于两个不同神经元,联系两个神经元的结构称为____________ 。在两个神经元之间传递信息的化学物质是____________ ,该物质以____________ 的形式释放到两神经元之间后,与后一神经元细胞膜上的_____________ 特异性结合,产生兴奋。
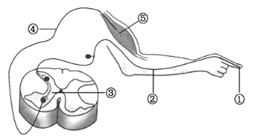
(2)图甲中反射弧通常由①、②、③、④
(3)在未受刺激时,②细胞膜两侧的电位处于
(4)图中的③、④属于两个不同神经元,联系两个神经元的结构称为
您最近一年使用:0次
2021-10-26更新
|
2189次组卷
|
29卷引用:广东省梅州市大埔县田家炳实验中学2021-2022学年高二上学期第一次段考(月考)生物试题
广东省梅州市大埔县田家炳实验中学2021-2022学年高二上学期第一次段考(月考)生物试题广东省高州市校际2021-2022学年高二11月联考生物试题广东省北师大珠海分校附属外国语学校2022-2023学年高二上学期期中生物试题(已下线)第2章 章末检测-2021-2022学年高二生物课后培优练(人教版选择性必修1)河北省衡水市武强中学2021-2022学年高二上学期第一次月考生物试题河北深州市长江中学2021-2022学年高二10月月考生物试题云南省昆明市官渡区一中2021-2022学年高二九月月考生物试题吉林省长春市北师大附属学校2021-2022学年高二上学期第一次月考生物试题江苏省常州市武进区礼嘉中学2021-2022学年高二上学期9月阶段教学质量调研生物(高考班)试题湖北省武汉市江夏实验高级中学2021-2022学年高二上学期10月考试生物试题河北省保定市顺平县中学2021-2022学年高二上学期第一次月考生物试题(已下线)第2章 神经调节 复习与提高(已下线)第一二章 综合测试卷-【上好课】2021-2022学年高二生物同步备课系列(浙科版2019选择性必修1)云南省文山州砚山县三中2021-2022学年高二下学期开学考试生物试题(已下线)考向17 神经调节-备战2023年高考生物一轮复习考点微专题(全国通用)浙江省湖州市吴兴高级中学2021-2022学年高二10月阶段性测试生物试题河南省南阳市2022-2023学年高二上学期第一次月考生物试题河南省新乡市原阳县三中2022-2023学年高二上学期第一次月考生物试题浙江省金华市东阳市三中2022-2023学年高二 9月月考生物试题河南省洛阳新学道高级中学2022-2023学年高二上学期第一次月考生物试题河南省驻马店市汝南县二中2022-2023学年-高二上学期第一次月考生物试题河北省邯郸市魏县五中2022-2023学年高二11月期中生物试题湖南省永州市宁远县明德湘南中学2022-2023学年高二上学期第一次月考生物试题湖南省郴州市永兴县童星学校2022-2023学年高三9月月考生物试题浙江省金华市江南中学2022-2023学年高二上学期10月阶段性考试生物试题(已下线)第05讲 神经冲动的产生和传导-【暑假自学课】2023年新高二生物暑假精品课(人教版2019选择性必修1)四川省雅安市三中2023-2024学年高二9月月考生物试题四川省宜宾市叙州区二中2023-2024学年高二10月月考生物试题云南省镇雄县第四中学2021-2022学年高二上学期期末考试生物试题
10 . 中暑常伴有发热症状,严重时还会损伤脏器。肾脏损伤时,血液中尿素氮等物质含量会升高,且肾组织切片检查会发现有不同程度的病理变化。为研究药物姜黄素预处理对不同程度中暑大鼠肾损伤的影响,所做实验如下表所示。请回答下列问题:
(1)调节体温的中枢位于______ ,在干热处理时大鼠主要通过______ 散热。
(2)乙组首次处理A为______ 。分别对每组大鼠进行不同时间干热处理的目的是______ 。
(3)本实验的自变量为______ 。
(4)表中实验结论B是______ 。
组别 项目 | 甲组 | 乙组 | ||||||
实验材料 | 相同生理状况的健康大鼠40只 | 相同生理状况的健康大鼠40只 | ||||||
首次处理 | 用适量200mg/kg姜苗连续灌胃天 | A | ||||||
再次处理(干热环境温度为41℃) | 均分为4组 | 均分为4组 | ||||||
干热 0min | 干热 50min | 干热 100min | 干热 150min | 干热 0min | 干热 50min | 干热 100min | 干热 150min | |
相应时间点检测大鼠血液中尿素氮含量,同时取肾组织制片,观察病理变化 | ||||||||
实验结果 |
| |||||||
实验结论 | B | |||||||

(1)调节体温的中枢位于
(2)乙组首次处理A为
(3)本实验的自变量为
(4)表中实验结论B是
您最近一年使用:0次
2019-12-08更新
|
176次组卷
|
2卷引用:2019年广东省湛江市高三一模理综生物试题


