1 . 大蒜是一种需要春化处理的植物,下表为二水早(小瓣紫皮蒜)和湘蒜(大瓣白皮蒜)在低温(2—6℃)处理不同天数后再播种得到的实验结果。回答下列问题:
播种前低温处理对大蒜出苗率的影响
注:播后出苗率=出苗数/播种蒜瓣数;
(1)由表中数据可见,一段时间的低温处理可__________ (填“促进”或“抑制”)大蒜的萌动。
(2)赤霉素具有促进种子__________ 的作用。某同学推测低温处理可能提高了大蒜内部的赤霉素水平,从而有利于打破大蒜的休眠状态。请选用合适的试剂进行实验来探究该同学的这个推测。
可选用试剂:赤霉素合成抑制剂、赤霉素
①将未萌动的二水早均分为甲、乙、丙三组,甲组常温下培养,_____________ 。经过适宜时间处理后,将各组蒜瓣播种,统计并计算播后出苗率。
②若实验结果为,出苗率________________________ ,则该同学的推测成立。
(3)研究发现,植物在接受春化作用后会产生春化素,且春化素可通过嫁接在同种植株间传导,诱导未春化的同种植株开花。此时,春化素作为一种__________ 信息,体现了生态系统中信息传递的作用是_________ 。
(4)大蒜的出苗率与品种、温度、光照和植株受伤害程度、基因的甲基化水平高低等因素有关,说明生物体表型与基因的关系为_______________ 。
播种前低温处理对大蒜出苗率的影响
品种 | 处理时间 | 播后出苗率% | ||
d | 10d | 15d | 25d | |
二水早 | 0 | 22.8 | 72.5 | 92.3 |
20 | 42.8 | 73.8 | 92.5 | |
30 | 64.3 | 76.8 | 92.8 | |
40 | 68.5 | 79.3 | 93.8 | |
湘蒜 | 0 | 22.5 | 72.8 | 92.3 |
20 | 42.3 | 75.8 | 92.5 | |
30 | 62.0 | 77.3 | 92.5 | |
40 | 68.3 | 78.8 | 94.5 | |
(1)由表中数据可见,一段时间的低温处理可
(2)赤霉素具有促进种子
可选用试剂:赤霉素合成抑制剂、赤霉素
①将未萌动的二水早均分为甲、乙、丙三组,甲组常温下培养,
②若实验结果为,出苗率
(3)研究发现,植物在接受春化作用后会产生春化素,且春化素可通过嫁接在同种植株间传导,诱导未春化的同种植株开花。此时,春化素作为一种
(4)大蒜的出苗率与品种、温度、光照和植株受伤害程度、基因的甲基化水平高低等因素有关,说明生物体表型与基因的关系为
您最近一年使用:0次
2 . 番茄果实发育受多种植物激素影响,市场上销售的植物激素或其类似物,如生长素类、乙烯利等物质,如果处理得当,对人体危害并不大,请回答下列问题:
(1)植物激素类似物属于植物生长调节剂,与植物激素相比,它具有_________ (至少写出2点)等优点。
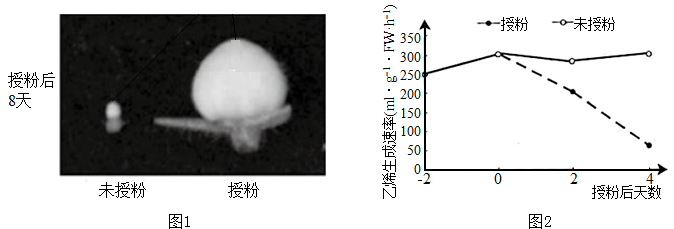
(2)研究者发现,未授粉及授粉8天后番茄的子房发育情况差异显著(如图1所示),结合生长素的生理作用推断,授粉后子房发育为果实的原因是_________ 。
(3)研究者检测授粉与未授粉番茄雌蕊产生乙烯的速率结果如图2所示。结合图1和图2推测乙烯对子房发育具有_________ 作用。
(4)为证实乙烯对子房发育的作用,研究者以野生型番茄为材料进行了如下表所示实验,请完善表格中的实验处理。表中I~Ⅲ处所需的试剂应分别选用_________(填选项前字母),实验结果支持上述推测。
(5)研究者经研究还发现,野生型番茄授粉后子房内赤霉素(GA)含量升高。为研究GA和乙烯对果实发育的影响,以野生型番茄和乙烯受体功能丧失的突变体S为材料进行实验,相关处理及结果如图3所示。
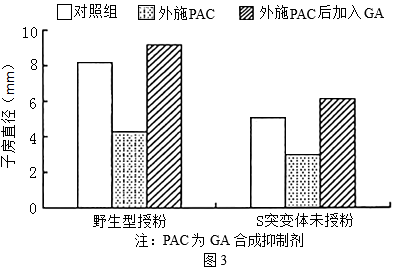
图中结果表明,GA的作用是_________ 。在未授粉情况下,突变体S比野生型体内GA合成相关基因表达量高,由此推测未授粉的S突变体子房可发育为果实的原因是_________ 。
(1)植物激素类似物属于植物生长调节剂,与植物激素相比,它具有
(2)研究者发现,未授粉及授粉8天后番茄的子房发育情况差异显著(如图1所示),结合生长素的生理作用推断,授粉后子房发育为果实的原因是
(3)研究者检测授粉与未授粉番茄雌蕊产生乙烯的速率结果如图2所示。结合图1和图2推测乙烯对子房发育具有
(4)为证实乙烯对子房发育的作用,研究者以野生型番茄为材料进行了如下表所示实验,请完善表格中的实验处理。表中I~Ⅲ处所需的试剂应分别选用_________(填选项前字母),实验结果支持上述推测。
授粉情况 | 授粉 | 未授粉 | 授粉 | 未授粉 |
实验处理 | 施加清水 | 施加Ⅰ | 施加Ⅱ | 施加Ⅲ |
子房是否发育为果实 | 是 | 否 | 否 | 是 |
| A.清水 | B.生长素 | C.生长素运输抑制剂 |
| D.乙烯利 | E.乙烯受体抑制剂 | |
图中结果表明,GA的作用是
您最近一年使用:0次
名校
3 . 根的生长受激素和营养物质的影响,科研人员以菊花为实验材料进行以下实验:
实验一:对菊花幼苗施用不同浓度的生长素,10天后对主根长度和侧根数目分别进行计数,结果如下表。
(1)生长素是对植物生长发育有重要_______ 作用的一类化合物,主要在_______________________________ 中合成。在成熟组织中,生长素可以通过输导组织进行___________ (填“极性”或“非极性”)运输。
(2)分析表中数据可知,促进侧根数量增加的生长素浓度___________ (填“促进”或“抑制”)主根伸长,生长素浓度为150×10-6mol/L时,__________ (填“促进”或“抑制”)侧根生长。由表中数据_______ (填“能”或“不能”)判断25×10-6mol/L的生长素对主根生长的影响.除生长素外,根部产生的_____________________ (激素)对植物的生长也具有促进作用。
(3)若生长素对菊花插条生根和侧根生长的作用效果相同,现有一未知浓度的生长素溶液,作用于插条后生根的数目是6,为确定该生长素的实际浓度,可将生长素溶液________ (处理)后作用于插条,若_________ ,则生长素浓度为150×10-6mol/L;若_________ ,则生长素浓度为50×10-6mol/L。
实验一:对菊花幼苗施用不同浓度的生长素,10天后对主根长度和侧根数目分别进行计数,结果如下表。
| 测定项目 | 施用生长素浓度(10-6mol/L) | ||||
| 0 | 50 | 100 | 150 | 200 | |
| 主根长度(相对值) | 1 | 0.9 | 0.7 | 0.5 | 0.3 |
| 侧根数目(个) | 4 | 6 | 8 | 6 | 3 |
(1)生长素是对植物生长发育有重要
(2)分析表中数据可知,促进侧根数量增加的生长素浓度
(3)若生长素对菊花插条生根和侧根生长的作用效果相同,现有一未知浓度的生长素溶液,作用于插条后生根的数目是6,为确定该生长素的实际浓度,可将生长素溶液
您最近一年使用:0次
2021-12-09更新
|
72次组卷
|
2卷引用:黑龙江省尚志市尚志中学2021-2022学年高二上学期期中生物试题
名校
4 . 低温会抑制烟草生长,并导致其开花提前,开花后养分向花蕾输送,造成烟叶的产量和品质下降。为探究植物激素脱落酸(ARA)对烟草抗低温和开花的影响,研究团队设置了4个组别(下表),其中CK表示正常温度,LT表示12℃低温处理。2周后,测定可溶性糖(小分子糖类)含量及开花情况,如果如下图和下表
1.上述实验应选取的烟草类型是_____ (单选)
2.烟草增加的可溶性糖的来源有_____ (编号选填)
①淀粉水解 ②糖酵解 ③光反应 ④碳反应
3.受低温胁追的植物细胞常表现为轻度的质壁分离状态。若使低温处理后的烟草液泡内可溶性糖的含量增加,则可以在显微镜下观察到_____,(单选)
4.结合上图和表,说明ABA对烟草抗低温和开花的影响:_____ 。
研究团队对相关基因的表达水平进行测定,结果如下图_____ (编号选填),而低温胁迫下ABA_____ (编号选填)
①加强TF4基因表达 ②加强FTS基因表达 ③加强SOCl基因表达
④减弱TF4基因表达 ⑤减弱FTS基因表达 ⑥减弱SOCl基因表达
6.为检测三种基因的表达量,请对下列操作进行排序_____ (编号排序)
①将RNA逆转录为DNA ②进行聚合酶链式反应
③从细胞样品中分离RNA ④检测分子浓度
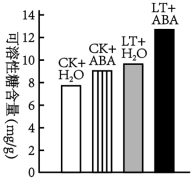
| 组别 | 开花情况 |
| CK+H2O CK+ABA LT+H2O LT+ABA | 50%烟株现蕾 无现蕾 全部现蕾,半数开花 花蕾较少,极少开花 |
1.上述实验应选取的烟草类型是_____ (单选)
| A.野外生长的低温敏感品种 | B.植物组织培养获得的低温敏感品种 |
| C.野外生长的抗低温品种 | D.植物组织培养获得的抗低温品种 |
①淀粉水解 ②糖酵解 ③光反应 ④碳反应
3.受低温胁追的植物细胞常表现为轻度的质壁分离状态。若使低温处理后的烟草液泡内可溶性糖的含量增加,则可以在显微镜下观察到_____,(单选)
| A.质壁分离程度增加 | B.细胞吸水涨破 |
| C.质壁分离复原 | D.无明显变化 |
研究团队对相关基因的表达水平进行测定,结果如下图
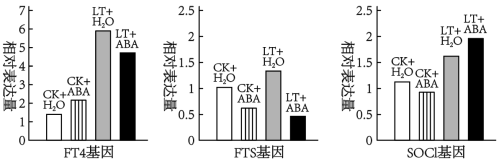
①加强TF4基因表达 ②加强FTS基因表达 ③加强SOCl基因表达
④减弱TF4基因表达 ⑤减弱FTS基因表达 ⑥减弱SOCl基因表达
6.为检测三种基因的表达量,请对下列操作进行排序
①将RNA逆转录为DNA ②进行聚合酶链式反应
③从细胞样品中分离RNA ④检测分子浓度
您最近一年使用:0次
2023-11-18更新
|
225次组卷
|
2卷引用:上海市复旦大学附中2023-2024学年高三上学期中生物试题
名校
5 . 干旱可诱导植物体内脱落酸(ABA)增加,以减少失水,但干旱促进ABA合成的机制尚不明确。研究者发现一种分泌型短肽(C)在此过程中起重要作用。科研人员分别用微量(0.1Μmol·L-1)的C或ABA处理拟南芥根部后,检测叶片气孔开度,结果如图1。为研究C的作用机理,科研人员又研究了 C基因缺失突变体在干旱处理中对N基因表达量的影响,已知N是催化ABA生物合成的关键酶,结果如图2。
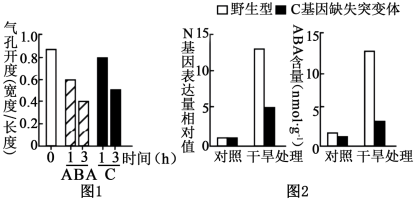
(1)分泌型短肽(C)合成、加工和分泌的过程依次经历的结构是_________ (选择序号并排序)
①内质网 ②线粒体 ③高尔基体 ④细胞核 ⑤质膜 ⑥核糖体
(2)在图1所示实验中,需要控制的无关变量是 。
(3)据图1结果,可以得知_________ (编号选填)
①C和ABA均可降低气孔开度,增加光合速率
②C和ABA可以影响植物的呼吸速率
③相对于ABA和C影响,时间对气孔影响更大
④C和ABA可以快速降低控制气孔开度相关基因的基因频率
(4)研究表明C可能通过促进N基因表达,进而促进ABA合成。图2中支持这 一结论的证据是__________________________ 。
(5)实验表明,野生型植物经干旱处理后,C在根中的表达远高于叶片;在根部外施的C可运输到叶片中。因此设想,干旱下根合成C运输到叶片促进N基因的表达。为验证此设想,进行了如下图和表所示的嫁接实验,干旱处理后,检测接穗叶片中C含量,又检测了其中N基因的表达水平。以接穗与砧木均为野生型的植株经干旱处理后的N基因表达 量为参照值,在表中填写假设成立时,与参照值相比N基因表达量的预期结果(用“远低于”“远高于”“相近”表示)。①____________ ;②____________ 。

(6)与野生型个体相比,C基因缺失突变体可能会出现的生理变化是
(1)分泌型短肽(C)合成、加工和分泌的过程依次经历的结构是
①内质网 ②线粒体 ③高尔基体 ④细胞核 ⑤质膜 ⑥核糖体
(2)在图1所示实验中,需要控制的无关变量是 。
| A.光照强度相同 | B.光合速率相同 | C.干旱程度相同 | D.气孔大小相同 |
(3)据图1结果,可以得知
①C和ABA均可降低气孔开度,增加光合速率
②C和ABA可以影响植物的呼吸速率
③相对于ABA和C影响,时间对气孔影响更大
④C和ABA可以快速降低控制气孔开度相关基因的基因频率
(4)研究表明C可能通过促进N基因表达,进而促进ABA合成。图2中支持这 一结论的证据是
(5)实验表明,野生型植物经干旱处理后,C在根中的表达远高于叶片;在根部外施的C可运输到叶片中。因此设想,干旱下根合成C运输到叶片促进N基因的表达。为验证此设想,进行了如下图和表所示的嫁接实验,干旱处理后,检测接穗叶片中C含量,又检测了其中N基因的表达水平。以接穗与砧木均为野生型的植株经干旱处理后的N基因表达 量为参照值,在表中填写假设成立时,与参照值相比N基因表达量的预期结果(用“远低于”“远高于”“相近”表示)。①
接穗 | 野生型 | 突变体 | 突变体 |
砧木 | 野生型 | 突变体 | 野生型 |
接穗叶片中N基因的表达量 | 参照值 | ① | ② |
(6)与野生型个体相比,C基因缺失突变体可能会出现的生理变化是
| A.生长素含量不同 | B.果实成熟的时间不同 |
| C.植物开花时间不同 | D.细胞分裂的方式不同 |
您最近一年使用:0次
名校
6 . 北方冬小麦生长期在少雨旱季,受旱灾影响的概率较高,其丰歉年景对国家粮食安全至关重要。某兴趣小组为探究不同程度的干旱条件对冬小麦光合速率和产量的影响,设置轻度干旱(W1)、中度干旱(W2)、重度干旱(W3)和对照(CK)四个处理的田间冬小麦试验,通过光合系统分析仪测定光合参数。下表1是不同条件下冬小麦光合参数的变化。
表1:不同条件下冬小麦光合参数的变化
根据上述材料,回答下列问题:
(1)对照组中,当光照强度等于冬小麦光补偿点时,叶肉细胞产生ATP的场所有__________________ 。
(2)研究发现,干旱条件下,冬小麦的细胞质中脯氨酸等可溶性小分子物质含量明显升高,其生物学意义是________________ 。
(3)干旱条件下,气孔导度的降低与_________ (植物激素)有关,在植物体中该激素还具有的功能是_______________________ 。
(4)小麦经济系数取决于小麦植株光合产物的积累、分配和转运,其籽粒产量的高低关键在于小麦干物质向籽粒的转运积累,为此兴趣小组进一步测定了冬小麦灌浆期叶片积累的有机物向地上部分Z1(除籽粒外)和地下部分Z2转运的分配率,如下表2。
表2:冬小麦干物质的分配率(%)
请根据表1、表2相关数据,分析干旱胁迫条件下,冬小麦籽粒干瘪,产量低的原因:______________ 。
表1:不同条件下冬小麦光合参数的变化
| 气孔导度[μmoLH2O/(m2·s)] | 叶绿素总量(mg·kg-1) | 冬小麦光补偿点[μmoL/(m2·s)] | 呼吸速率[μmoLCO2/(m2·s)] |
CK | 0.2601 | 1.02 | 53.60 | 0.9363 |
W1 | 0.0957 | 0.84 | 54.18 | 1.1533 |
W2 | 0.0754 | 0.57 | 55.89 | 1.4790 |
W3 | 0.0453 | 0.46 | 56.40 | 1.7594 |

根据上述材料,回答下列问题:
(1)对照组中,当光照强度等于冬小麦光补偿点时,叶肉细胞产生ATP的场所有
(2)研究发现,干旱条件下,冬小麦的细胞质中脯氨酸等可溶性小分子物质含量明显升高,其生物学意义是
(3)干旱条件下,气孔导度的降低与
(4)小麦经济系数取决于小麦植株光合产物的积累、分配和转运,其籽粒产量的高低关键在于小麦干物质向籽粒的转运积累,为此兴趣小组进一步测定了冬小麦灌浆期叶片积累的有机物向地上部分Z1(除籽粒外)和地下部分Z2转运的分配率,如下表2。
表2:冬小麦干物质的分配率(%)
处理 | Z1(除籽粒外) | Z2 |
CK | 30.84 | 18.32 |
W1 | 32.15 | 16.31 |
W2 | 31.65 | 26.09 |
W3 | 30.48 | 33.81 |
请根据表1、表2相关数据,分析干旱胁迫条件下,冬小麦籽粒干瘪,产量低的原因:
您最近一年使用:0次
7 . 黄瓜属于浅根性蔬菜作物,对盐渍、高温、干旱等逆境条件适应性较差。为探讨在干旱胁迫下,钙与脱落酸对黄瓜幼苗光合作用的影响,科学家将幼苗分成A、B、C三组,A组正常栽培;B、C组分别用脱落酸(ABA)、CaCl2+ABA喷施叶片处理后,在干旱胁迫条件下栽培,一周后测定了相关指标,结果如下表。回答下列问题:
注:SOD(超氧化物歧化酶)具有抗氧化作用,可清除植物在不利环境下产生的自由基等。
(1)内源脱落酸是一种植物激素,其作用是促进叶和果实的衰老和脱落、_____________ (至少答出一点)等。
(2)据表分析,本实验中A组作为___________ 组;B、C组黄瓜幼苗的净光合速率明显低于A组的原因是干旱胁迫使___________ 下降,导致暗反应所需的CO2不足;胁迫下的黄瓜幼苗通过增加细胞中游离氨基酸的含量来提高_______________ ,以吸收更多土壤中的水分来抵抗干旱胁迫。
(3)已知Ca2+在植物抗逆过程中能防止膜脂过氧化,具有稳定膜结构的作用,请从表找出支持该结论的证据:________________ ,使清除自由基的效果更明显。
(4)进一步研究发现:干旱胁迫一周后与处理前比较,植物的地下部分与地上部分干重的比值明显提高,其意义是________________ 以抵御干旱胁迫带来的伤害。
(5)本实验叶面喷施ABA的浓度是1.0mg·L-1,为了进一步探究提高干旱胁迫下黄瓜幼苗净光合速率的最适ABA浓度,请你写出实验设计思路____________________ 。
分组 | 气孔导度(mmol·m-2·s-1) | 净光合速率(μmolCO2·m-2·s-1) | 游离氨基酸(μg·g-1) | SOD活性(U·mg-1pro) |
A组 | 0.54 | 14.5 | 1.83 | 16.1 |
B组 | 0.34 | 8.0 | 2.48 | 13.8 |
C组 | 0.39 | 9.0 | 2.16 | 14.5 |
注:SOD(超氧化物歧化酶)具有抗氧化作用,可清除植物在不利环境下产生的自由基等。
(1)内源脱落酸是一种植物激素,其作用是促进叶和果实的衰老和脱落、
(2)据表分析,本实验中A组作为
(3)已知Ca2+在植物抗逆过程中能防止膜脂过氧化,具有稳定膜结构的作用,请从表找出支持该结论的证据:
(4)进一步研究发现:干旱胁迫一周后与处理前比较,植物的地下部分与地上部分干重的比值明显提高,其意义是
(5)本实验叶面喷施ABA的浓度是1.0mg·L-1,为了进一步探究提高干旱胁迫下黄瓜幼苗净光合速率的最适ABA浓度,请你写出实验设计思路
您最近一年使用:0次
8 . 某同学利用不同浓度生长素(IAA)和赤霉素(GA3)对某番茄品种(S-47)进行试验,来研究植物激素对种子萌发的影响;下表为相关实验数据,其中发芽率(%)=(发芽种子数/供试种子数)×100%。请回答下列问题:
(1)生长素的主要合成部位是______ ;其在胚芽鞘的运输______ (填“会”或“不会”)消耗能量;本实验结果______ (填“能”或“不能”)体现IAA对番茄种子(S-47)萌发作用的两重性,理由是______ 。
(2)由表可知,随着GA3浓度的升高,番茄种子(S-47)的发芽率逐渐______ ,种子萌发的调控过程中,与GA3作用效果相反的植物激素是______ 。
(3)本实验的实验结论不一定适用于其他植物,原因是______ 。
| 生长调节刻种类 | 浓度(mg/L) | 发芽率(%) |
| 对照组 | 蒸馏水 | 80 |
| IAA | 2 | 96.13 |
| 4 | 93.13 | |
| 6 | 89.46 | |
| 8 | 84.06 | |
| 10 | 81.33 | |
| GA3 | 100 | 97.93 |
| 200 | 94.2 | |
| 300 | 91.6 | |
| 400 | 88.4 | |
| 500 | 81 |
(1)生长素的主要合成部位是
(2)由表可知,随着GA3浓度的升高,番茄种子(S-47)的发芽率逐渐
(3)本实验的实验结论不一定适用于其他植物,原因是
您最近一年使用:0次
9 . 为研究遮阴对棉花植株光合作用特性和叶绿素含量的影响,研究人员设置了如下实验:CK、CK′选用中棉19号,1、1′和2、2′分别选用“春矮早”和“9418”棉花品种,在自然光照、遮阴(50%自然光强)条件分别进行实验,实验结果如下表。回答下列问题:
(1)光反应阶段,棉叶吸收的太阳光能主要储存在______ 中。暗反应的场所是______ 。
(2)与对照组相比,自然光照下,实验组棉叶中的叶绿素含量______ (填“较高”或“较低”)。叶肉细胞中,叶绿素主要吸收_____ 光。
(3)三个品种的棉花植株中,光合速率最大的是_______ 。除光合速率指标外,其他指标也支持上述结论,表现为_______ 。
(4)相同条件下,三个棉花品种的光合特性不同,本质上是由______ 决定的。干旱条件下,喷洒适宜浓度的ABA有利于棉花植株抗旱,原理是_______ 。
| 处理 | 光合速率(μmol/m2·s) | 胞间CO2浓度(μmol/mol) | 气孔阻力相对值 | 叶绿素含量(mg/dm2) | |
| 正常 | CK | 16.95 | 284.4 | 0.33 | 4.4 |
| 1 | 28.20 | 280.9 | 0.27 | 4.1 | |
| 2 | 16.37 | 280.6 | 0.46 | 4.5 | |
| 遮阴 | CK′ | 17.13 | 273.6 | 0.54 | 3.0 |
| 1′ | 20.38 | 273.5 | 0.39 | 3.4 | |
| 2′ | 16.95 | 273.3 | 0.51 | 3.5 | |
(2)与对照组相比,自然光照下,实验组棉叶中的叶绿素含量
(3)三个品种的棉花植株中,光合速率最大的是
(4)相同条件下,三个棉花品种的光合特性不同,本质上是由
您最近一年使用:0次
解题方法
10 . 马铃薯块茎达到生理成熟后,会进入很长一段时间的休眠期。处于休眠过程中的块茎仍维持着最基础的生理功能。只有经过一定时间的贮藏后,块茎才能解除休眠并进一步发芽生长。植物激素是影响马铃薯块茎休眠和芽萌发的重要因素,下表是马铃薯块茎休眠过程中主要内源激素的含量变化。请回答下列问题:
马铃薯块茎休眠过程中主要内源激素的含量变化
(1)生长素主要促进________ 的分裂,可与细胞分裂素协同促进细胞分裂的完成。赤霉素在促进细胞分裂时与表中的________ 作用相抗衡。
(2)马铃薯块茎的休眠过程,不是由单一激素起调节作用,而是_________ 。
(3)据表可知,________ 是可以促进马铃薯块茎打破休眠的植物激素。研究人员用乙烯拮抗剂处理马铃薯块茎,结果块茎提前发芽,因此推断出乙烯有利于____________ (填“块茎休眠”或“芽萌发”)的启动和维持。
(4)研究表明马铃薯休眠时期的长短主要与品种的遗传特性有关,同时也会受到环境、栽培措施和贮藏条件等因素的影响。综上可知,马铃薯块茎休眠和芽萌发是由__________ 共同完成的。
马铃薯块茎休眠过程中主要内源激素的含量变化
| 休眠时期 | 含量 | ||||
| 生长素 | 赤霉素 | 细胞分裂素 | 脱落酸 | 生长素 | |
| 休眠初期 | 低水平 | 低水平 | 低水平 | 低水平 | 高水平 |
| 休眠深期 | 高水平 | 低水平 | 低水平 | 高水平 | 低水平 |
| 休眠释放期(萌发前) | 低水平 | 高水平 | 高水平 | 低水平 | 低水平 |
(2)马铃薯块茎的休眠过程,不是由单一激素起调节作用,而是
(3)据表可知,
(4)研究表明马铃薯休眠时期的长短主要与品种的遗传特性有关,同时也会受到环境、栽培措施和贮藏条件等因素的影响。综上可知,马铃薯块茎休眠和芽萌发是由
您最近一年使用:0次


